REVIEW: Correlation between Bioeffector Characteristics of Sphingolipids and the Structure of Their Hydrophobic Fragment
E. V. Dyatlovitskaya
Shemyakin and Ovchinnikov Institute of Bioorganic Chemistry, Russian Academy of Sciences, ul. Miklukho-Maklaya 16/10, Moscow GSP-7, 117871 Russia; fax: (095) 335-7103; E-mail: jgmol@ibch.siobc.ras.ru
Submitted June 9, 1997
The effect of the structure of the hydrocarbon sphingoid base chain (length, presence of double bonds, and steric configuration of functional groups) and the length of the fatty acid residue of a ceramide molecule on the bioregulatory activities of sphingoids and ceramides is reviewed. Particular emphasis is placed on the role of functional groups (OH- and NH2-) typical of the sphingoid chain.
KEY WORDS: bioeffector, sphingenine, stereospecificity, chemical structure, ceramide, sphingolipid
Abbreviations: C2, C6, C8, etc.) the number of C-atoms in the hydrocarbon chain of sphingoids and fatty acids; C2-ceramide) N-acetylsphingenine; N-acylsphinganine) dihydroceramide.
In terms of chemical structure and functional activity, sphingolipids
belong to one of the most diverse classes of lipids. These compounds
were given such a name for their enigmatic properties (from Greek
"sphinx") [1]. A large number of studies
have revealed that sphingolipids exert a significant effect on the
structure of biological membranes and lipoproteins [2, 3]; anchor some membrane
proteins [4, 5]; contribute to
the processes of cell recognition, growth, and differentiation; mediate
reception of hormones, toxins, growth factors, and cytotoxins; and have
immunomodulation activities (see reviews [6-19]).
Biological activities of large glycosphingolipids have been studied for many years. The main emphasis of these studies was placed on the correlation between the biological effect of the molecule and structure of its polar fragment (hydrocarbon chain). The contribution of ceramide, the hydrophobic fragment of sphingolipid, was not taken into consideration.
However, in the last decade the structure of hydrophobic fragments of sphingolipids--N-acylsphingosines (ceramides) and sphingoid base--has attracted considerable attention because it was shown that a free sphingoid sphingenine inhibits protein kinase C in vitro and has a significant effect on the cell response associated with activation of protein kinase C in thrombocytes [20], neutrophils [21], and myeloid leukemic cells HL-60 [22]. It should be noted that the chemical structure of sphingoid bases and their N-acyl-derivatives significantly affects their biological efficiency, modification of different parts of the molecules having different biological effects.
The goal of this work was to review the presently available data on the correlation between structural modification of hydrophobic fragments of sphingolipids and their biological efficiency.
STRUCTURE OF SPHINGOLIPIDS
Sphingolipids is a collective name of hundreds of lipid compounds that share a common feature (sphingoid base), but have differing chemical structures and biological functions. Sphingoid bases are amino alcohols with a long hydrocarbon chain. Sphingenine, an aminodiol with a chain of 18 carbon atoms and a double trans-bond in position 4,5, is the most abundant sphingoid base in sphingolipids of human and animal cells (Fig. 1).
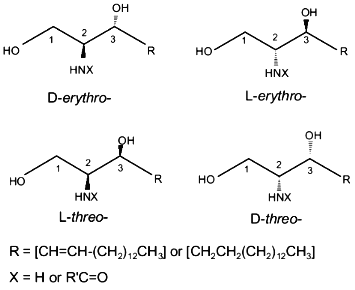
The sphingenine molecule has two asymmetric carbon atoms (positions 2 and 3). Therefore, it can attain four stereoisomeric configurations with different spatial positions of functional groups (D-erythro-, D-threo-, L-erythro-, and L-threo-) (Fig. 2). Natural sphingolipids contain the D-erythro-isomer of sphingenine.Fig. 1. Structure of sphingolipids.
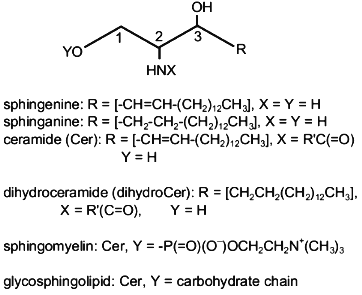
There are three functional groups in a sphingenine molecule: two OH-groups and one NH2-group. Modification of these groups produces different types of sphingolipids. For example, N-acylation of the NH2-group by fatty acids produces ceramides (Fig. 1), which are precursors of more complicated sphingolipids (sphingomyelin and glycosphingolipids) (Fig. 1). Therefore, any sphingolipid molecule contains a hydrophobic fragment (one or two hydrocarbon chains with functional groups).Fig. 2. Stereoisomers of sphingenine.
ROLE OF SPHINGOID STRUCTURE
As mentioned above, the D-erythro-isomer of sphingenine is the dominant sphingoid base of natural sphingolipids. Dihydrosphingenine (D-erythro-sphinganine) and phytosphingosine (4-hydroxy-D-erythro-sphinganine) are found as minor components.
Hydrocarbon chain. The biological activity of sphingoid bases was shown to depend significantly of their chemical structure. It is quite obvious that interaction of functional groups of a sphingoid with a cell targets plays a dominant role in its biological activity. However, the capacity for such interaction significantly depends on the hydrocarbon chain structure. It was shown that 18 carbon atoms is the optimal chain length of a sphingoid for inhibiting protein kinase C. Sphingoids containing shorter (C11-C16) or longer (C20) hydrocarbon chains exert lower inhibitory effects [23]. Similar dependence on the sphingoid base length was observed for the inhibition of differentiation of HL-60 cells induced by phorbol 12-myristate-13-acetate and inhibition of growth of the CHO cells [23].
The configuration of a double bond also plays a significant part in the biological activity. For example, the replacement of trans-isomer of D-erythro-sphingenine by its cis-form causes an approximately twofold decrease in the ability to inhibit protein kinase C in vitro [23], but increases DNA synthesis in Swiss 3T3 cells [24]. On the other hand, both cis and trans isomers of D-erythro-sphingenine were found to have almost the same capabilities for releasing Ca2+ from intracellular stores [24].
Double bond saturation (i.e., conversion of sphingenine into sphinganine) also has a significant effect on the biological activity of the molecule. It has been shown that low concentrations (1 µM) of the racemic mixture of DL-erythro-sphinganine, a metabolic precursor of natural sphingenine, and DL-threo-sphinganine, a stereoisomer of DL-erythro-sphinganine, stimulates proliferation of Swiss 3T3 cells more effectively than sphingenine [25]. High concentrations of sphinganine (15-20 µM) exert only an insignificant effect on this process [24, 25]. Both D-erythro-sphinganine and L-erythro-sphinganine were less effective stimulators of protein kinase C in the cytosol of Jurkat T-cells than D-erythro-sphingenine or L-erythro-sphingenine [26]. On the other hand, exogenous sphingenine and sphinganine exerted virtually the same effect on apoptosis of HL-60 cells [27].
Functional groups. Steric configuration of functional groups plays a decisive part in the regulation of sphingenine activities. As mentioned above, the sphingenine molecule has two asymmetric carbon atoms (at positions 2 and 3) and can attain four stereoisomeric configurations (D-erythro-, D-threo-, L-erythro-, and L-threo-) (Fig. 2). Natural sphingolipids contain only the D-erythro-isomer of sphingenine.
Studies of artificial sphingenines showed that steric configuration in some cases has a considerable effect on biological activity. For example, DL-threo-sphinganine and L-threo-sphingenine exert higher toxic effect on Swiss 3T3 cells than the corresponding erythro-isomers [25]. Only D-erythro-isomers of sphinganine and sphingenine in low concentrations stimulate proliferation of Swiss 3T3 fibroblasts, whereas DL-threo-sphinganine or L-threo-sphingenine are inactive [24]. Only D-erythro-sphingenine is active in the reaction of Ca2+ release from intracellular stores [24]. However, different sphingenine stereoisomers have virtually the same effects on apoptosis of HL-60 cells [27]. Similarly, no stereospecificity of the sphingenine isomers was found in the inhibition of protein kinase C in vitro [23].
What is the role of individual functional groups of sphingenine?
As mentioned above, there are three functional groups in a sphingenine molecule: two OH-groups (positions 1 and 3) and one NH2-group (position 2) (Fig. 1). Experiments in vitro revealed that protein kinase C is inhibited not only by sphingenine [20, 28], but also by lysoglycosphingolipids (lyso-GM2, sulfogalactosylsphingenine, and galactosylsphingenine) [28, 29], i.e., sphingenine derivatives where the OH-group in position 1 is replaced by a carbohydrate chain (Fig. 1). In contrast to sulfogalactosylsphingenine, the corresponding glycosphingolipids do not induce inhibition of protein kinase C in vitro. The kinetic curves of protein kinase C inhibition by lysosphingolipids and by sphingenine are similar [28]. Therefore, the OH-group at position 1 is not involved in the protein kinase C inhibition. On the other hand, in contrast to sphingenine, lysosphingolipids (glycosylsphingenines) were found to be ineffective in the regulation of phosphorylation of epidermal growth factor (EGF) receptor in A431 cells [30] or hormone binding to GH3 cells [31]. Therefore, these reactions require the presence of a free hydroxyl group in position 1.
If the hydroxyl group in position 1 is replaced by a phosphate group, the resulting molecule (sphingenine-1-phosphate), like sphingenine itself, is a second messenger [17]. Because only one form of sphingenine-1-phosphate (natural) has been described in the literature, it is not considered in this review. It should only be noted that substitution of the hydroxyl group in position 1 by a phosphate group produces a biologically effective sphingolipid.
The amino group apparently plays a key role in the biological effects of sphingolipids. As noted above, sphingenine is the most common natural sphingoid. Only insignificant amounts of sphingenine were found in the free state. N,N-Dimethylsphingenine, a compound with a higher positive charge, was found in brain tissues in addition to sphingenine [32]. Natural N,N-dimethylsphingenine is a more effective inhibitor of purified protein kinase C in A431 cells than sphingenine, the activity of the compound depending significantly on its steric configuration. D-Erythro-isomer is a more effective inhibitor than L-erythro-, D-threo-, or L-threo-isomers [33]. However, according to other authors [23], N,N-dimethylsphingenine is a less effective inhibitor of protein kinase C than sphingenine. Moreover, it was shown that N-methyl,N-isopropylsphingenine, another acyl-derivative of sphingenine, activates rather than inhibits protein kinase C [23]. It was also shown that N,N-dimethylsphingenine is a significantly more effective stimulator of phosphorylation of the EGF receptor than sphingenine or N-methylsphingenine [34]. In addition, sphingenine derivatives demonstrated differing effects on neurite growth efficiency in NS-20Y murine neuroblastoma cells. N,N-Dimethylsphingenine was found to be ten times more effective as an inhibitor of neurite growth than sphingenine [35]. N,N,N-Trimethylsphingenine inhibits protein kinase C even more effectively [36]. Both N,N-dimethylsphingenine and N,N,N-trimethylsphingenine inhibit tumor cell growth in vitro more effectively than sphingenine [37]. However, both sphingenine and its N-substituted derivatives have virtually the same effect on the functions of neutrophils [38].
Thus, the existing literature shows that a chain of 18 carbon atoms, a trans double bond in position 4,5, and an NH2-group are required to give the optimal biological efficiency of sphingoids. The structures of natural sphingenine or lysosphingolipids meet these requirements. However, some types of biological activities additionally require a free hydroxyl group to be present at position 1, which is observed only in the sphingenine molecule.
Usually, replacement of hydrogen atoms of the NH2-group by alkyl substituents increase the biological activity of sphingenine. This effect can be attributed to increased positive charge of the molecule. However, in some cases such a replacement causes a decrease in the biological activity of sphingenine, which is probably due to the fact that alkylated sphingoids are not involved in metabolic reactions.
Although steric selectivity is not observed universally among sphingenines, D-erythro-isomer (natural sphingenine) is the most effective bioregulator.
ROLE OF CERAMIDE STRUCTURE
As shown in the previous section, biological activity of sphingoids depends on their structure. Acylation of the NH2-group of sphingenine by fatty acids produces ceramides (Fig. 1), which have been found in cells either as free molecules or as fragments of complex sphingolipids. It has been found during the last decade that free ceramides are very important second messengers and mediators of cell growth stimulation [25, 39] or inhibition [40-50] (including cell cycle arrest at stage G0/G1 [43, 46, 49]); induction of differentiation [40, 42, 48, 50, 51], senescence [52], and apoptosis [27, 45, 47, 49, 53-62]. Ceramides also regulate evolution of H2O2 by activated neutrophils [63, 64], stimulate HIV production by infected HL-60 cells [65], contribute to immune reactions [18], and have a notable part in the biosynthesis of prostaglandins [66, 67] and steroids [67].
It has been shown that all biological functions of ceramides depend on their chemical structure. As with sphingoids (see above), the biological activity of ceramides depends on the sphingoid base structure. However, ceramide is an N-acyl-derivative of sphingoid. Therefore, there are two hydrocarbon chains in the ceramide molecule (sphingoid base and fatty acid). Each of the two chains can contribute to the biological activity of the molecule.
Sphingoids. As mentioned above, optimal bioregulation activity is observed in D-erythro-sphingenine (natural sphingoid). As a result, biological activity of ceramides was tested using synthetic analogs of natural sphingenine. However, ceramides based on artificial sphingoids have also been tested by some authors. For example, studies of the role of the steric configuration of ceramide sphingoids in the regulation of cell proliferation revealed that the four steric isomers of synthetic N-acetylsphingenine (D-erythro-, D-threo-, L-erythro-, and L-threo-) readily penetrate the cell membrane and inhibit the growth of HL-60 cells [45]. In contrast, steric isomers of N-acetylsphinganine (dihydroceramide) exerted significantly differing effects on the proliferation of these cells: D-erythro- and L-erythro-isomers had virtually no effect, whereas D-threo- and L-threo-isomers strongly inhibited cell growth [45]. A certain steric specificity was also observed in the effects of ceramides on protein phosphatase 2A from T9 cells of rat glioma or rat brain [68]. Although all steric isomers of N-acetylsphingenine cause a significant activation of this enzyme, the effect induced by the D-erythro-isomer is noticeably higher [69]. In contrast to this, the racemic mixture of N-acetylsphinganine isomers without a trans double bond in position 4,5 of the sphingoid chain induced inhibition of protein phosphatase activity [55, 69]. Rather interesting results were obtained in studies of the effect of steric isomers of N-octanoylsphingenine on apoptosis induction in the U937 cells [60]. Threo-isomers of this ceramide were shown to be approximately seven times more active stimulators of DNA fragmentation than erythro-isomers. DL-Erythro-N-octanoylsphinganine also induced apoptosis, although to a lesser extent than D-erythro-N-octanoylsphingenine [60]. Therefore, rather scarce information on the effects of steric isomerization of ceramide sphingoids on their biological activity does not provide clear evidence of an advantage of natural configuration (D-erythro-), because in some cases threo-isomers were found to be more active [60].
However, it is beyond doubt that the trans double bond in position 4,5 of a sphingoid plays a significant part in the biological activities of the molecule. In contrast to synthetic ceramides which easily penetrate cell membranes, their dihydro-derivatives (dihydroceramides) have no effect on various biological processes. For example, N-acetyl-, N-hexanoyl-, and N-octanoylsphinganines did not inhibit cell growth [43, 45, 49, 51, 52], did not stimulate cell differentiation [49, 51], did not induce dephosphorylation of Rb (a protein product of the retinoblastoma gene) [43], and did not induce apoptosis [27, 45, 49, 53, 58, 59, 62]. However, some authors reported that not only ceramides, but also dihydroceramides are able to stimulate apoptosis [54, 60]. It was also shown that not only dihydroceramides with short acyl chains, but also dihydroceramides with long fatty acid residues do not affect cell differentiation [51]. It was found in studies of mechanisms of interaction between ceramides and cell targets that in contrast to ceramides, corresponding dihydroceramides inhibit rather than stimulate activity of protein phosphatases [55, 69] and do not change the activity of an isoform of protein kinase C [57] and phospholipase D [70] (which are inhibited by ceramides). On the other hand, both N-acetylsphingenine and N-acetylsphinganine were shown to inhibit activity of acyl-CoA-cholesterol acyltransferase in hamster ovary cells [71].
Thus, in most cases the presence of a double trans-bond in position 4,5 of the sphingoid chain of ceramides is a necessary prerequisite for their biological activity.
Fatty acids. Most studies of biological activity of ceramides were performed with short acyl chain (C2, C6, and C8) synthetic analogs of natural compounds because they easily penetrate the cell membrane. However, the metabolic activity of these compounds differs considerably from natural ceramides, the activity being dependent on the fatty acid chain length. For example, N-acetylsphingenine and N-acetylsphinganine are virtually not involved in metabolic reactions at all [40, 45, 71], whereas N-octanoylsphingenine is rapidly incorporated into cellular sphingolipids [72]. On the other hand, hydrocarbon chains of sphingoid and fatty acid facilitate the interaction between ceramides and cell targets. Therefore, the length of the acyl chain should have an effect on this interaction.
The literature shows that the ceramide-mediated effects on biological processes in cells indeed depend on the length of the fatty acid chain of the ceramide molecule. For example, in contrast to short-chain (C2, C6) ceramides, C18-ceramide does not inhibit proliferation of lymphocytes [47]. Natural ceramides are less effective inducers of cell differentiation in neuroblastoma than short-chain ceramides [51]. C18-Ceramide does not induce DNA fragmentation in U937-cells [47], does not stimulate protein phosphatase activity [69], and does not inhibit phospholipase D [70] and acyl-CoA-cholesterol acyltransferase [71]. However, some authors have reported that apoptosis of neurons is stimulated by natural ceramides and N-acetylceramide to approximately the same extent [58]. DNA fragmentation in U937 cells treated with natural ceramides was found to be even higher than in cells treated with C2-ceramide [56]. It was shown that the presence of hydroxy acids in natural ceramides decreases significantly apoptosis induction [56]. It should be noted that the efficiency of artificial ceramides also depends on the fatty acid chain length. For example, N-octanoylsphingenine is more active than N-acetyl- or N-hexanoylsphingenines in promotion of differentiation of the DJM-1 cells (a line of ceratinocytes) [48]. The ceramide-induced activation of protein phosphatase also depends on the acyl chain length: at a concentration of 1 µM, N-dodecanoylsphingenine and N-myristoylsphingenine demonstrate maximal activities, whereas N-acetylsphingenine is significantly less active, and N-octanoylsphingenine is absolutely inactive [69].
What is the mechanism of the biological activity of fatty acid residues of ceramides?
It is absolutely evident that functional groups of ceramides (hydroxy and amide) are involved in their interaction with cell targets. Long hydrocarbon chains of sphingoid and fatty acid facilitate this interaction by making the molecular conformation more favorable. It is also obvious that molecular conformation depends on the hydrophobic interaction between the hydrocarbon chains of a sphingoid and fatty acid, therefore, on the sphingoid chain structure (length, presence or absence of double bond) and the fatty acid chain structure (length, presence or absence of double bonds, their number and location, presence of hydroxyl groups). Short-chain acyls are virtually incapable of inducing such interactions. The efficiency of hydrophobic interaction rises with increasing fatty acid chain length. Hydrophobic interaction between hydrocarbon chains of sphingoids and fatty acids in ceramides can even reduce the difference between the biological activities of ceramides and dihydroceramides. For example, natural ceramides isolated from benign ovarian tumors were found to have substantially different composition of fatty acids and sphingoid bases than natural ceramides isolated from malignant ovarian tumors. In addition to sphingenine, typical of ceramides of normal tissue of human ovary, the ceramides isolated from malignant and benign tumors were abundant with sphinganine (51 and 27% in malignant and benign tissue, respectively). However, in spite of such a significant difference in the content of ceramides and dihydroceramides, these ceramide pools had approximately the same activities with respect to the inhibition of proliferation of the CaOv cells (human ovary carcinoma cells) [73]. The hydrophobic interaction between hydrocarbon chains in C18-ceramide may explain the fact that this compound does not cause inhibition of the proliferation of lymphocytes [47], induction of DNA fragmentation in U937 cells [47], stimulation of protein phosphatase activity [69], and inhibition of the phospholipase D [70] and acyl-CoA-cholesterol acyltransferase activities [71] (in contrast to short-chain ceramides).
Our analysis of the whole body of experimental data on the biological activities of sphingoid bases and ceramides showed that steric factors contribute to these activities only in a few cases. However, because natural sphingolipids contain D-erythro-isomer of sphingenine, the data on the steric specificity of the biological activity of a sphingoid bases is only of purely theoretical significance. The problem of variation of the regulation activity of sphingoid bases induced by metabolic modification of their functional groups appears more interesting and important for practical purposes.
Comparison between biological activities of sphingoids and ceramides with short acyl chains allows the contribution of amine and amide groups to be determined. It should be noted that in contrast to a sphingenine molecule, stearylamine, a sphingenine analog that does not contain hydroxy groups, in most cases does not affect cell processes: it did not change the protein kinase C activity [26], caused no mitogenic effect on Swiss 3T3 cells [74] and did not modulate H2O2 evolution by neutrophils [63]. Therefore, it can be concluded that all functional groups of sphingoids and ceramides are required to cause their biological activity. However, because stearylamine is able to inhibit DNA synthesis in T-cells [44] and growth of CHO cells [23], the presence of only an amino group is sufficient to cause some types of biological activities. Alkylation of an amino group of sphingenine modifies its biological activity [75] and may even cause an opposite activity [23, 33]. Acylation of an amino group (i.e., sphingenine conversion into ceramide) may cause the disappearance [23] of the biological activity or change its sign. For example, cell differentiation is inhibited by sphingenine [23], but it is activated by ceramides [40, 42, 48, 50, 51]. Certain types of biomodulation activity of sphingenine (stimulation of apoptosis and inhibition of cell growth) coincide with those in ceramides, although the extent of their manifestation is different. It is interesting to note that N-octylsphingenine stimulates apoptosis more effectively than N-octanoylsphingenine, and some authors even suggested that the -C=O group is not necessary to stimulate apoptosis [60].
It is rather difficult to conclude from the available data whether or not the hydroxyl groups of sphingoids and ceramides play a significant role in the biological activities of these molecules. It is evident that hydroxyl groups are involved in the interaction of the second messengers (sphingoids and ceramides) with cell targets. It is also known that in contrast to ceramide, GlcCer (a glycosylated derivative of ceramide, where the hydroxyl group in position 1 of the hydrocarbon chain of the sphingoid is replaced by glucose) stimulates rather than inhibits cell proliferation [76]. Also in contrast to ceramide, an N-acetylsphingenine-containing ganglioside GM3 does not affect DNA biosynthesis in HL-60 cells [47]. It can be suggested that introduction of a complex polar group (carbohydrate chain) to position 1 of the sphingoid chain modifies its interaction with cell targets, and the bioregulatory effects of glycosphingolipids (including gangliosides) are caused by other mechanisms [47, 77].
The literature contains no information on the role of a hydroxyl group at position 3 of a sphingoid chain of ceramide on its biological activity. It was only reported that sphingomyelinase fails to decompose a sphingomyelin analog containing no hydroxyl group at position 3 of the sphingoid chain of a ceramide fragment [78].
Thus, our survey of the literature on the biological activities of sphingoid and ceramides, the core structures of sphingolipids, showed that their chemical structure plays a significant role in the biological effects. Because these compounds are the second messengers of external signal transduction to cell, and because they regulate growth, differentiation, and apoptosis of cells, it is not inconceivable that some of these natural metabolites (e.g., N,N-dimethylsphingenine) or their synthetic analogs [37] can be used in antitumor therapy.
REFERENCES
1.Thudichum, J. L. W. (1962) A Treatise on the
Chemical Constituents of the Brain (a Facsimile Edition of the
Original), Bailliere, London.
2.Barenholz, Y., and Gatt, S. (1982), in
Phospholipids (Hawthorne, J. N., and Ansell, G. B., eds.)
Elsevier, Amsterdam, pp. 129-177.
3.Barenholz, Y., and Thompson, T. E. (1980),
Biochim. Biophys. Acta, 604, 129-158.
4.Lederkremer, R. V., Lima, C., Ramirez, M. I., and
Casal, O. L. (1990) Eur. J. Biochem., 192, 337-345.
5.Takeda, A. (1993) Adv. Lipid Res.,
26, 293-317.
6.Fishman, P. H., and Brady, R. O. (1976)
Science, 194, 906-915.
7.Hakomori, S. (1981) Ann. Rev. Biochem.,
50, 733-764.
8.Hakomori, S. (1984) Trends Biochem. Sci.,
9, 453-455.
9.Curratolo, W. (1987) Biochim. Biophys. Acta,
906, 137-160.
10.Sweeley, C. C. (1991) in Biochemistry of
Lipids, Lipoproteins, and Membranes (Vance, D. E., and Vance, J.
E., eds.) Elsevier, Amsterdam, pp. 327-361.
11.Dyatlovitskaya, E. V., and Bergelson, L. D.
(1987) Biochim. Biophys. Acta, 907, 125-143.
12.Hannun, Y. A., and Bell, R. M. (1989)
Science, 243, 500-507.
13.Merrill, A. H., Jr. (1991) J. Bioenerg.
Biomembr., 23, 83-104.
14.Merrill, A. H., Jr., Hannun, Y. A., and Bell, R.
M. (1993) Adv. Lipid Res., 25, 1-24.
15.Hakomori, S., and Igarashi, Y. (1995) J.
Biochem., 118, 1091-1103.
16.Hannun, Y. A. (1996) Handbook Lipid Res.,
8, 177-204.
17.Spiegel, S., and Merrill, A. H., Jr. (1996)
FASEB J., 10, 1388-1397.
18.Ballou, L. R., Laulederkind, S. J. F., Rosloniec,
E. F., and Raghow, R. (1996) Biochim. Biophys. Acta,
1301, 273-287.
19.Shayman, J. A. (1996) L. Amer. Soc.
Nephrol., 7, 171-182.
20.Hannun, Y. A., Loomis, C. R., Merrill, A. H.,
Jr., and Bell, R. M. (1986) J. Biol. Chem., 261,
12604-12609.
21.Wilson, E., Olcott, M. C., Merrill, A. H., Jr.,
and Bell, R. M. (1986) J. Biol. Chem., 261,
12616-12623.
22.Merrill, A. H., Jr., Sereni, A. M., Stevens, V.
L., Hannun, Y. A., Bell, R. M., and Kinkade, J. M., Jr. (1986) J.
Biol. Chem., 261, 12610-12615.
23.Merrill, A. H., Jr., Nimkar, S., Menaldino, D.,
Hannun, Y. A., Loomis, C. R., Bell, R. M., Tyagi, S. R., Lambeth, J.
D., Stevens, V. L., Hunter, R., and Liotta, D. C. (1989)
Biochemistry, 28, 3138-3145.
24.Olivera, A., Zhang, H., Carlson, R. O., Mattie,
M. E., Schmidt, R. R., and Spiegel, S. (1994) J. Biol. Chem.,
269, 17924-17930.
25.Hauser, J. M. L., Buehrer, B. M., and Bell, R. M.
(1994) J. Biol. Chem., 269, 6803-6809.
26.Pushkareva, M. Yu., Bielawska, A., Menaldiv, D.,
Liotta, D., and Hannun, Y. A. (1993) Biochem. J., 294,
699-703.
27.Jarvis, W. D., Fernari, F. A., Jr., Traylor, R.
S., Martin, H. A., Kramer, L. B., Erukulla, R. K., Bittman, R., and
Grants, S. (1996) J. Biol. Chem., 271, 8275-8284.
28.Hannun, Y. A., and Bell, R. M. (1989) Clin.
Chim. Acta, 185, 333-346.
29.Hannun, Y. A., and Bell, R. M. (1987)
Science, 235, 670-674.
30.Faucher, M., Girones, N., Hannun, Y. A., Bell, R.
M., and Davis, R. J. (1988) J. Biol. Chem., 263,
5319-5327.
31.Winicov, I., and Gershengorn, M. C. (1988) J.
Biol. Chem., 263, 12179-12182.
32.Nudelman, E. D., Levery, S. V., Igarashi, Y., and
Hakomori, S. (1992) J. Biol. Chem., 267,
11007-11016.
33.Igarashi, Y., Hakomori, S., Toyukuni, T., Dean,
B., Fujita, S., Sugimoto, M., Ogawa, T., El-Chendy, K., and Racker, E.
(1989) Biochemistry, 28, 6796-6800.
34.Igarashi, Y., Kitamura, K., Toyukuni, T., Dean,
B., Fenderson, B., Ogawa, T., and Hakomori, S. (1990) J. Biol.
Chem., 265, 5385-5389.
35.Uemura, K., Hara, A., and Taketomi, T. (1993)
J. Biochem., 114, 610-614.
36.Hakomori, S., and Igarashi, Y. (1993) Adv.
Lipid Res., 25, 147-162.
37.Endo, K., Igarashi, Y., Nisar, M., Zhou, Q., and
Hakomori, S. (1991) Cancer Res., 51, 1613-1618.
38.Kimura, S., Kawa, S., Ruan, F., Nisar, M.,
Sadahira, Y., Hakomori, S., and Igarashi, Y. (1992) Biochem.
Pharmacol., 44, 1585-1595.
39.Olivera, A., Buckley, N. E., and Spiegel, S.
(1992) J. Biol. Chem., 267, 26121-26127.
40.Okazaki, T., Bielawska, A., Bell, R. M., and
Hannun, Y. A. (1990) J. Biol. Chem., 265,
15823-15831.
41.Dbaibo, G., Obeid, L. M., and Hannun, Y. A.
(1993) J. Biol. Chem., 268, 17762-17766.
42.Dobrowsky, R. T., Werner, M. H., Castellino, A.
M., Chao, M. V., and Hannun, Y. A. (1994) Science, 265,
1596-1599.
43.Dbaibo, G., Pushkareva, M. Yu., Jayadev, S.,
Schwarz, J. K., Horowiz, J. M., Obeid, L. M., and Hannun, Y. A. (1995)
Proc. Natl. Acad. Sci. USA, 92, 1347-1351.
44.Borchardt, R. A., Lee, W. T., Kalen, A., Buckley,
R. H., Peters, C., Schiff, S., and Bell, R. M. (1994) Biochim.
Biophys. Acta, 1212, 327-336.
45.Bielawska, A., Crane, H. M., Liotta, D., Obeid,
L. M., and Hannun, Y. A. (1993) J. Biol. Chem., 268,
26226-26232.
46.Kuroki, J., Hirokawa, M., Kitabayashi, A., Lee,
M., Horiuchi, T., Kawabata, Y., and Miura, A. B. (1996)
Leukemia, 10, 1950-1958.
47.Olshefski, R., Taylor, B., Heitger, A., Hasegawa,
A., and Ladisch, S. (1996) Eur. J. Biochem., 241,
47-55.
48.Wakita, H., Tokura, Y., Yagi, H., Nishimura, K.,
Furukawa, F., and Takigawa, H. (1994) Arch. Dermatol. Res.,
286, 350-354.
49.Jayadev, S., Liu, B., Bielawska, A., Lee, J. Y.,
Nazaire, F., Pushkareva, M. Yu., Obeid, L. M., and Hannun, Y. A. (1995)
J. Biol. Chem., 270, 2047-2052.
50.Bielawska, A., Linardic, C. M., and Hannun, Y. A.
(1992) FEBS Lett., 307, 211-214.
51.Riboni, L., Prinetti, A., Bassi, R., Caminiti,
A., and Tettamanti, G. (1995) J. Biol. Chem., 270,
26868-26875.
52.Venable, M. E., Lee, J. Y., Smyth, M. J.,
Bielawska, A., and Obeid, L. M. (1995) J. Biol. Chem.,
270, 30701-30708.
53.Obeid, L. M., Linardic, C. M., Karolak, L. A.,
and Hannun, Y. A. (1993) Science, 259, 1769-1771.
54.Jarvis, W. D., Kolesnick, R. N., Fornari, F. A.,
Traylor, R. S., Gewirtz, D. A., and Grant, S. (1994) Proc. Natl.
Acad. Sci. USA, 91, 73-77.
55.Wolff, R. A., Dobrowsky, R. T., Bielawska, A.,
Obeid, L. M., and Hannun, Y. A. (1994) J. Biol. Chem.,
269, 19605-19609.
56.Ji, L., Zhang, G., Uematsu, S., Akahori, Y., and
Hirabayashi, Y. (1995) FEBS Lett., 358, 211-214.
57.Lee, J. Y., Hannun, Y. A., and Obeid, L. M.
(1996) J. Biol. Chem., 271, 13169-13174.
58.Brugg, B., Michel, P. P., Agid, Y., and Ruberg,
M. (1996) J. Neurochem., 66, 733-739.
59.Kaipia, A., Chun, S.-Y., Eisenhauer, K., and
Hsueh, A. J. W. (1996) Endocrinology, 137,
4864-4870.
60.Karasavvas, N., Erukulla, R. K., Bittman, R.,
Lokshin, R., and Zakeri, Z. (1996) Eur. J. Biochem.,
236, 729-737.
61.Wiesner, D. A., and Dawson, G. (1996)
Glycoconjug. J., 13, 327-333.
62.Hartfield, P. J., Mayne, G. C., and Murray, A. W.
(1997) FEBS Lett., 401, 148-152.
63.Naramura, T., Abe, A., Balazovich, K. J., Wu, D.,
Suchard, J., Boxer, L. A., and Shayman, J. A. (1994) J. Biol.
Chem., 269, 18384-18389.
64.Wong, K., Li, X. B., and Hanchuk, N. (1995) J.
Biol. Chem., 270, 3056-3062.
65.Rivas, C. I., Golde, W. B., Vera, J. C., and
Kolesnick, R. N. (1994) Blood, 83, 2191-2197.
66.Ballou, L. R., Chao, C. P., Holness, M. A.,
Barker, S. C., and Raghow, R. (1992) J. Biol. Chem.,
267, 20044-20050.
67.Santana, P., Llanes, L., Hernandez, I.,
Gonzalez-Robayana, I., Tabraue, C., Gonzalez-Reyes, J., Quintana, J.,
Estevez, F., Ruiz de Galarreta, C. M., and Fanjul, L. F. (1996)
Endocrinology, 137, 2480-2489.
68.Dobrowsky, R. T., and Hannun, Y. A. (1992) J.
Biol. Chem., 267, 5048-5051.
69.Dobrowsky, R. T., Kamibayashi, C., Mumby, M. C.,
and Hannun, Y. A. (1993) J. Biol. Chem., 268,
15523-15530.
70.Venable, M. E., Bielawska, A., and Obeid, L. M.
(1996) J. Biol. Chem., 271, 24800-24805.
71.Ridgway, N. D. (1995) Biochim. Biophys.
Acta, 1256, 39-46.
72.Abe, A., Wu, D., Shayman, J. A., and Radin, N. S.
(1992) Eur. J. Biochem., 210, 765-773.
73.Rylova, S. N., Kozlov, A. M., Kogtev, L. S.,
Gaenko, G. P., and Dyatlovitskaya, E. V. (1997) Biochemistry
(Moscow), 62, 1053-1056.
74.Zhang, H., Bucley, N. E., Gilson, K., and
Spiegel, S. (1990) J. Biol. Chem., 265, 76-81.
75.Ohta, H., Yatomi, Y., Sweeney, E. A., Hakomori,
S., and Igarashi, Y. (1994) FEBS Lett., 355,
267-270.
76.Marsh, N. L., Elias, P. M., and Holleran, W. M.
(1995) J. Clin. Invest., 95, 2903-2909.
77.Nagai, Y. (1995) Behav. Brain. Res.,
66, 99-104.
78.Lister, M. D., Ruan, Z. S., and Bittman, R.
(1995) Biochim. Biophys. Acta, 1256, 25-30.