Proton Transfer through the Membrane-Water Interfaces in Uncoupled Mitochondria
V. I. Yurkov*, M. S. Fadeeva, and L. S. Yaguzhinsky
Belozersky Institute of Physico-Chemical Biology, Lomonosov Moscow State University, 119992 Moscow, Russia; fax: (7-095) 939-3181; E-mail: yurkovv@yandex.ru* To whom correspondence should be addressed.
Received September 21, 2004
Increase in maximal respiration rate of uncoupled mitochondria in response to increase in concentration of non-penetrating buffer has been demonstrated. This phenomenon did not depend on chemical structure of uncouplers and composition of the non-penetrating buffer. Use of covalently attached pH probe, FITC, revealed that at low buffer concentration (3 mM) the H+-pump functioning results in local increase in proton concentration on the outer surface of the inner mitochondrial membranes. In other words, local H+ gradient was generated. Increase in buffer concentration up to 20 mM caused sharp decrease in this gradient, which occurred in parallel to increase in the respiration rate. It is concluded that both effects described here may be attributed to the process of proton transfer through the interfaces of the mitochondrial membrane: the rate of respiratory H+ pumps of uncoupled mitochondria under conditions of low buffer capacity of medium is limited by the stage of proton release from outer surface of the coupling membrane. The inhibition mechanism of respiration by high concentrations of uncouplers is also discussed.
KEY WORDS: mitochondria, local H+ gradients, uncouplers, proton transport
Transmembrane proton transport through mitochondrial and artificial phospholipid membranes was studied in details due to its key role during transformation of energy of oxidative reactions in the systems of oxidative phosphorylation in mitochondria and chloroplasts. Significant contributions to our knowledge on functioning of these systems have been made by V. P. Skulachev. Using a representative group of aromatic acids, he provided experimental explanation of principle of the uncoupling effect of uncouplers [1]. In collaboration with E. A. Liberman (and using penetrating ions), Skulachev demonstrated the existence of electric potential on the inner mitochondrial membrane [2]. The present work represents a logical continuation of studies underlying action mechanisms of uncouplers. It is based on the concept of the existence of local H+ gradients on the inner mitochondrial membrane under conditions of respiratory chain functioning [3-5].
It was usually considered that hydrophobic layer of phospholipid membrane plays a role of kinetic barrier in this process. However, results of experiments on various model systems provided certain evidence that during transmembrane proton transfer the stage of H+ transfer through the membrane-water interphase may also have a relatively high kinetic barrier. The existence of such a barrier was originally demonstrated by Drachev, Kaulen, and Skulachev [6] using models of rhodopsin plaques. Later Heberle et al. confirmed their results [7]. This effect was also observed in other systems [8]. On the planar membrane, we also registered local H+ gradient at the interphase; formation of this gradient was determined by high kinetic barrier in the reaction of proton release from a surface of lipid bilayer [9]. Recently, we have demonstrated formation of local H+ gradients in zones of interphases on the mitochondrial membranes [3] under phosphorylating conditions. The goal of the present study was to demonstrate existence of kinetic barriers in the reactions of proton transfer through the interphase of the inner mitochondrial membranes in uncoupled mitochondria and to demonstrate the effect of the kinetic barriers on the respiration rate.
MATERIALS AND METHODS
Rat liver mitochondria were isolated as described in [3].
The main measurements were carried out using freshly isolated mitochondria within 1-2.5 h after their isolation. Mitochondrial respiration was registered by using a Clark type electrode and LP-7E polarograph. Protein was determined by the biuret method.
Method of registration of local gradients of H+ activity on the external surface of the inner mitochondrial membrane. Since FITC (fluorescein isothiocyanate) can cross the inner mitochondrial membrane, covalent linkage of the probe occurs at both sides (external and internal) of the membrane [3, 10]. Spectral changes of the probe appearing during H+ pump functioning were registered by a differential scheme using a Hitachi-557 spectrophotometer. Spectral changes at the internal and external probe fractions were superimposed, and for their separate registration parallel measurements at low (3 mM) and high (20 mM) concentrations of weakly membrane-penetrating buffer (Hepes) were carried out. In the latter case the value of external H+ gradient was minimal. So it is reasonable to accept that all changes can be attributed to changes of absorbance of the internal probe fraction. Comparison of results obtained at these buffer concentrations indicates the existence of H+ gradient. This gradient appears on the outer surface of the inner mitochondrial membrane in media with low buffer concentration. In all experiments, pH of the incubation medium was constant (7.5). The method of covalent linkage of FITC and registration of local H+ gradients in phosphorylating mitochondria was described in detail [1].
The following chemicals were used in this study: Hepes, Tris, Mes, dimethyl sulfoxide (Amresco, USA); KCl, MgSO4, fluorescein isothiocyanate (Fluka, Switzerland); EDTA, sucrose (Merck, Germany); fatty acid free albumin (ICN, Germany); TTFB, ADP, K2HPO4, succinate, rotenone, TTFA, myxothiazol, phenol, pentachlorophenol, trichlorophenol, FCCP (Sigma, USA).
RESULTS
At the first stage of this study, we measured changes in respiration rate of uncoupled mitochondria in response to increase in buffer concentrations in the incubation medium. Several uncouplers of different chemical structure were used. These included three phenolic compounds (phenol (I), trichlorophenol (II), and pentachlorophenol (III)), benzimidazole derivative (tetrachlorotrifluoromethyl benzimidazole (TTFB) (IV)), and phenylhydrazone derivative (carbonyl cyanide p-(trifluoromethoxy)phenylhydrazone (FCCP) (V)).
In all cases, the rate of uncoupled respiration increased with the increase in buffer concentration (Figs. 1 and 2). Figures show that changes in buffer concentration as such (in the absence of uncouplers) did not influence the respiration rate V4 (state 4 according to Chance). The effect of the increase in uncoupled respiration induced by increase in buffer capacity of the incubation medium was observed only in mitochondria characterized by high respiratory control of 3-6; this effect was also observed within 1-2 h after mitochondria isolation. Longer storage of mitochondrial preparations at 0-4°C usually caused a decrease in the respiratory control. Mitochondria entered a new functional state characterized by other effects [4].
Fig. 1. Effect of increasing concentrations of weakly penetrating buffer in the incubation medium on the mitochondrial respiration rate. Respiration medium contained 2 mM succinate, 1 µM rotenone, pH 7.5; uncouplers: a) phenol; b) trichlorophenol; c) FCCP; d) pentachlorophenol. Broken and solid lines show changes in the respiration rate at 3 and 20 mM Hepes, respectively. The incubation medium contained 0.25 mM EDTA, 1 mM magnesium sulfate, and 10 mM KCl. Sucrose was added to adjust medium osmolarity to 120 mOsm.
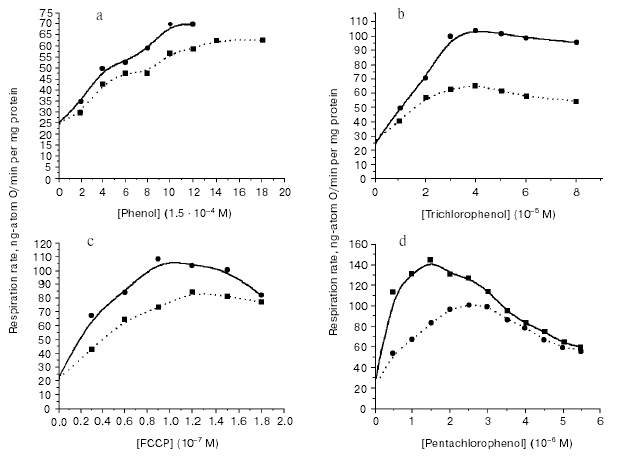
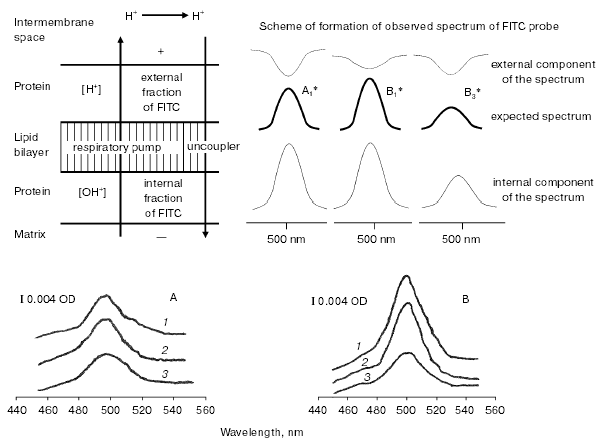
Using highly effective uncouplers (III, IV) (Figs. 1d and 2b), it was convenient to observe pronounced effect of respiration inhibition by high concentrations of these compounds; this also eliminated dependence of the respiration rate on buffer capacity of the medium. In the second stage of this study, we registered local increase in hydrogen ion activity on the outer surface of the inner mitochondrial membrane in uncoupled mitochondria. Since the discussed problem is rather important, it is relevant to describe here the scheme of experiment in more detail (Fig. 3). The experiment was carried out using uniformly FITC labeled mitochondria under conditions when ~30% of the bound probe was in non-dissociated form. According to the scheme, oxidation of substrate of respiratory H+ pumps (succinate) in the experimental cuvette results in increase in matrix pH, dissociation of FITC, and increased absorbance of the matrix fraction of the probe. Figure 3 shows expected spectrum (A1*, B1* (intern.)). If this occurs simultaneously with local increase in H+ (externally) due to H+ release by the respiratory pump, the external probe should be partially converted into non-dissociated form and cause an extinction decrease in the external FITC-fraction (Fig. 3, A1*, B1* (extern.)). Superimposition of spectra of external and internal probe fractions yields the expected spectrum shown at Fig. 3 (A1* (exp.)). According to the theory [11] and model experiments [9], non-penetrating buffer accelerates H+ transfer through the interphase. Increase in non-penetrating buffer concentration should result in decrease in the external local H+ gradient (Fig. 3, cf. A1* and B1* (extern.)); so the expected spectrum will be more intensive than in weak buffer (Fig. 3, cf. A1* and B1* (exp.)). Sharp increase in uncoupler concentration should result in increase in relative H+ influx into matrix and decrease in matrix pH. Sharp decrease in intensity of absorbance of matrix probe fraction should result in sharp decrease in the expected spectrum (Fig. 3, cf. B1* and B3* (exp.)). Results of experiments (Fig. 3, A and B) interpreted in accordance with this scheme revealed local decrease in H+ activity of the outer surface of the mitochondrial membrane, which can be attributed to H+ pump functioning in the uncoupled mitochondria. For example, at 10-6 and 2*10-6 M concentrations of TTFB accelerating mitochondrial respiration, intensity of observed spectra of FITC probe in dilute buffer (3 mM) was significantly lower (Fig. 3, A, curves 1, 2) than in the presence of higher concentration (20 mM) of the non-penetrating buffer (Fig. 3, B, curves 1, 2). At inhibitory concentration of uncoupler (5*10-6 M TTFB), there was sharp decrease in intensity of probe spectrum attributed to decrease in matrix pH (Fig. 3, B, curve 3).Fig. 2. Effect of weakly penetrating buffers of different chemical structures on the mitochondrial respiration rate. a) 3 mM Mes (broken line), 20 mM Mes (solid line); b) 3 mM Hepes (broken line), 20 mM Hepes (solid line). Experimental conditions were the same as described in Fig. 1.
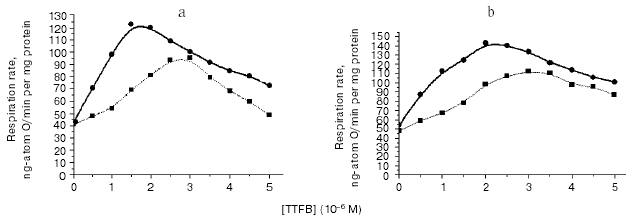
Fig. 3. Scheme of H+ transport and scheme of expected changes in spectra of pH probe, which should appear during H+-pump functioning in uniformly FITC-labeled mitochondria under different experimental conditions. The expected spectra were obtained as the result of overlay of spectra of external and internal fractions of the FITC probe. A1*) low concentration of the non-penetrating buffer, low concentration of uncoupler; B1*) high concentration of non-penetrating buffer, low concentration of uncoupler; B3*) high concentration of the non-penetrating buffer, high concentration of uncoupler; A, B) differential spectra of FITC-labeled mitochondria obtained at low (3 mM) and high (20 mM) concentrations of the non-penetrating buffer, respectively. Succinate (2 mM) and rotenone (1 µM), pH 7.5, were added to both cuvettes; tenoyltrifluoroacetone (TTFA) (1 µM) was added to the control cuvette. Curves 1, 2, and 3 were obtained with TTFB added (in both cuvettes) at 1, 3, and 6 µM concentration, respectively.
DISCUSSION
In this study, we have demonstrated the phenomenon of increase in mitochondrial respiration in response to an increase in non-penetrating buffer concentration in the mitochondrial incubation medium. This phenomenon did not depend on chemical nature of uncouplers (I-V) (see Figs. 1 and 2). This phenomenon was not particularly related to chemical structure of molecules of buffer (Fig. 2). These data suggest that this phenomenon does not represent an artifact and it may be attributed to actual uncoupling effect of compounds I-V. We will discuss the nature of this phenomenon below.
The rates of respiration and transmembrane proton flux are related to each other. The latter may be limited by the stage of proton release from the interphase. This was demonstrated using model system (planar lipid bilayer) [9], non-phosphorylating mitochondria [4], and also phosphorylating mitochondrial [3] and submitochondrial particles [5]. Experiments [12] and theoretical calculations [11] indicate that low molecular weight buffer should accelerate processes of proton attachment/detachment on polyelectrolytes and, consequently, accelerate reaction of proton detachment from the interphase in the cases when membrane surface includes acid-base groups.
Measurements have demonstrated that the rate of respiratory H+ pump functioning is limited by the stage of H+ release from the membrane; this results in formation of a local gradient on the outer surface of the inner mitochondrial membrane with excessive concentration of protons (Fig. 3, A and B, curves 1, 2). Figure 3 shows that increase in Hepes concentration in the medium caused sharp decrease in H+ gradient; this occurred in parallel with the increase in the uncoupled respiration rate. The following evident chain of events may be proposed. Increase in concentration of a non-penetrating buffer accelerates the slow stage of proton detachment from the membrane surface. This exhausts local H+ gradient on the membrane surface; transmembrane flux of protons increases and respiration rate increases as well (Figs. 1 and 2). During sharp increase in uncoupler concentration, relative rate of H+ transfer into matrix becomes greater than the effective rate constant of proton efflux from mitochondria. This results in appearance of a steady state characterized by lower matrix pH (Fig. 3, B, curve 3), respiration rate decreases, and sensitivity of respiration to non-penetrating buffer almost disappears (Figs. 1d and 2b). The latter observation suggests “translocation” of the rate-limiting stage of the process from the outer side of the interphase to the inner one. These results suggest that decrease in matrix pH is the direct reason responsible for the earlier described phenomenon [1] of reduction of mitochondrial respiration rate by uncouplers.
In conclusion, it should be noted that inward and outward channels of proton pumps are located within interphase zones [13]. In fact, these interphases play a role of proton harvesting and proton dissipating antennas. So in this study the rate-limiting stages of functioning of respiratory H+ pumps and their regulation are considered.
REFERENCES
1.Skulachev, V. P. (1969) Energy Accumulation in
the Cell [in Russian], Nauka, Moscow.
2.Skulachev, V. P. (1972) Energy Transformation in
Biomembranes [in Russian], Nauka, Moscow.
3.Solodovnikova, I. M., Yurkov, V. I., Ton'shin, A.
A., and Yaguzhinskii, L. S. (2004) Biofizika, 49,
47-56.
4.Kozlova, M. A., Gramadsky, K. B., Solodovnikova, I.
P., Krasinskaya, I. P., Vinogradov, A. V., and Yaguzhinsky, L. S.
(2003) Biofizika, 48, 443-453.
5.Yaguzhinsky, L. S., Lapin, M. V., and Krasinskaya,
I. P. (1998) FEBS Lett., 440, 223-225.
6.Drachev, L. A., Kaulen, A. D., and Skulachev, V. P.
(1984) FEBS Lett., 178, 331-335.
7.Heberle, J., Riesle, J., Thiedemann, G., Osterhelt,
D., and Dencher, N. A. (1994) Nature, 370, 379-382.
8.Prats, M., Tocanne, J. F., and Tessier, J. (1985)
Eur. J. Biochem., 149, 663-668.
9.Antonenko, Yu. N., Koubasnjuk, O. N., and
Yaguzhinsky, L. S. (1993) Biochim. Biophys. Acta, 1150,
45-50.
10.Krasinskaya, I. P., Korshunov, S. S., Kachanov,
O. Yu., and Yaguzhinsky, L. S. (1997) Biochemistry (Moscow),
62, 364-370.
11.Junge, W., and McLauglin, S. (1987) Biochim.
Biophys. Acta, 890, 1-5.
12.Tanford, W. (1965) Physical Chemistry of
Biopolymers [Russian translation], Nauka, Moscow.
13.Wikstrom, M., Verkhovsky, M., and Hummer, G.
(2003) Biochim. Biophys. Acta, 1604, 61-65.