Interaction of Yeast Mitochondria with Fatty Acids and Mitochondria-Targeted Lipophilic Cations
E. I. Sukhanova, T. A. Trendeleva, and R. A. Zvyagilskaya*
Bach Institute of Biochemistry, Russian Academy of Sciences, Leninsky pr. 33, 119071 Moscow, Russia; fax: (495) 954-2732; E-mail: renata_z@inbi.ras.ru* To whom correspondence should be addressed.
Received November 5, 2009
The effect of fatty acids and mitochondria-targeted lipophilic cations (SkQ1, SkQ3, MitoQ, and C12TPP) on tightly-coupled mitochondria from yeasts Dipodascus (Endomyces) magnusii and Yarrowia lipolytica was investigated. Micromolar concentrations of saturated and unsaturated fatty acids were found to decrease the membrane potential, which was recovered almost totally by ATP and BSA. At low, micromolar concentrations, mitochondria-targeted lipophilic cations are “relatively weak, mild uncouplers”, at higher concentrations they inhibit respiration in state 3, and at much higher concentrations they induce swelling of mitochondria, possibly due to their prooxidant and detergent action. At very low, not uncoupling concentrations, mitochondria-targeted lipophilic cations profoundly promote (potentiate) the uncoupling effect of fatty acids. It is conceivable that the observed uncoupling effect of lipophilic cations can be, at least partially, due to their interactions with the endogenous pool of fatty acids.
KEY WORDS: yeast, Yarrowia lipolytica, Dipodascus (Endomyces) magnusii, mitochondria, fatty acids, SkQ1, SkQ3, MitoQ, C12TPP, respiration, transmembrane potential, swellingDOI: 10.1134/S000297910020033
Abbreviations: CCCP, carbonyl cyanide m-chlorophenylhydrazone; CTAB, cetyltrimethylammonium bromide; C12TPP, dodecyltriphenylphosphonium; MitoQ, 10-(6′-ubiquinonyl)decyltriphenylphosphonium; ROS, reactive oxygen species; SkQ1, 10-(6′-plastoquinonyl)decyltriphenylphosphonium; SkQ3, 10-(5-methylplastoquinonyl)decyltriphenylphosphonium; Δψ, mitochondrial transmembrane potential.
There are more than 300 theories of aging known to date [1]. One of the most popular modern theories is still
the free radical theory of aging. According to this theory, aging and
age-related pathologies are crucially dependent on oxidative stress
caused by reactive oxygen species (ROS) [2].
Indeed, much evidence shows that aging is accompanied by increasing
accumulation of oxidative changes in DNA, especially in mitochondrial
DNA (mDNA), proteins, and lipids [3]. Inasmuch as
mDNA encodes proteins of the mitochondrial electron transport chain and
of the ATP-synthase complex, the age-related oxidative stress should
result in substantial impairments of energy supply, which, in turn,
should lead to senescence. Besides, ROS frequently induce cell death
(apoptosis or necrosis).
Therefore, it is not surprising, that attempts are constantly being made to delay senescence using antioxidants [3]. The most promising in this respect was the use of MitoQ (10-(6′-ubiquinonyl)decyltriphenylphosphonium) – the positively charged (and hence, transporting predominantly, if not exclusively, into mitochondria) lipophilic cation triphenylphosphonium bonded by a C10-aliphatic chain with ubiquinone, a component of the mitochondrial respiratory chain with antioxidant activity (see review [4] and references therein).
Mitochondria-targeted lipophilic antioxidants offer advantages over conventional water-soluble antioxidants as they are transported into and accumulated within cells and mitochondria in conformity with the membrane potential generated on the cytoplasmic or mitochondrial membrane, respectively. As a result, their concentrations in mitochondria would increase several orders of magnitude in comparison with the initial low, nontoxic nanomolar and sub-nanomolar concentrations. Moreover, lipophilic antioxidants, once accumulated in the lipid bilayer of the inner mitochondrial membrane, can be reduced (regenerated) by components of the respiratory chain, which ensures their repetitive functions.
V. P. Skulachev (see reviews [3, 5] and references therein) proposed replacing ubiquinone in the molecule of MitoQ by the potentially more powerful natural antioxidant plastoquinone normally functioning in the photosynthetic electron-transport chain (in chloroplasts) under severe conditions of elevated oxygen concentrations and ROS production. Within the framework of a project supervised by V. P. Skulachev, a series of plastoquinone derivatives (referred to collectively as SkQ, where Sk means “penetrating cation”, “Skulachev ion” – a term put forward by D. Green [6], and Q is plastoquinone) were synthesized [3, 5, 7]. Cationic derivatives of plastoquinone were found to easily penetrate through bilayer lipid membranes forming a diffusion-potential of the expected value [3, 7]. They are accumulated in cell mitochondria, thus protecting these organelles from ROS-induced apoptosis or necrosis [3, 7], decreasing lipid peroxidation and protein carbonylation in aging animals [3, 8], increasing median lifespan of podospora, daphnia, drosophila, and mice [3, 9], and retarding the development of various age-related diseases such as heart and kidney infarctions, heart arrhythmia, brain stroke [10], some types of tumors [11], and ocular diseases (cataract, glaucoma, retinopathies, uveitis, and others) [8]. Interestingly, in some cases the antioxidant concentration “window” (at which the antioxidant activity dominated over the prooxidant one) for SkQ was significantly wider than that for MitoQ, just as predicted by Skulachev.
Elucidating of mechanisms underlying the effect of SkQ derivatives on entire organisms is hampered by the complexity of these objects. Mitochondria are still the simplest while adequate model for such researches. Tightly-coupled yeast mitochondria have some advantages over animal mitochondria as they display diminished rate of endogenous respiration (which permits investigation of the oxidation of individual substrates) and, as we showed [12], lack a Ca2+/Pi-dependent pore (which facilitates interpretation of the data obtained). Using tightly-coupled mitochondria from yeasts Yarrowia lipolytica and Dipodascus magnusii, we showed that mitochondrially-targeted lipophilic cations with delocalized charge (SkQ1, SkQ3, MitoQ, and C12TPP) can act, depending on the concentrations used, as antioxidants, uncouplers, prooxidants, and detergents. Low concentrations of SkQ1, SkQ3, MitoQ, and C12TPP potentiated the uncoupling effect of long-chain unsaturated fatty acids.
MATERIALS AND METHODS
Reagents. ADP, ATP, UDP, GTP, GDP, pyruvate, malate, succinate, palmitate, fatty-acid-free BSA, alamethicin, atractyloside, safranine O, carboxyatractyloside, carbonyl cyanide m-chlorophenylhydrazone (CCCP), EDTA, EGTA, Tris, sorbitol, and mannitol were purchased from Sigma-Aldrich (USA); Coomassie G-250 and Zymolyase 20T were from MP Biomedicals (USA); ETH129 was from Fluka (Germany); yeast extract was from Difco (USA). Other reagents of the highest quality were from domestic suppliers.
Organisms and cultivation conditions. In this work we used yeasts D. magnusii strain VKM 261 and Y. lipolytica. The Y. lipolytica strain was isolated as a pure isolate from epiphytic microflora of salt-excreting arid plants (the central Negev Highlands of Israel) and identified on the basis of its morphological, biochemical/physiological characteristics, and of quantitative chemotaxonomic and molecular marker analyses as Yarrowia lipolytica (Wick.) van der Walt & Arx. [13]. Yeast cells were cultivated at 28°C in agitated (220 rpm) 750-ml Erlenmeyer flasks in 100 ml of a semi-synthetic medium [14] containing 1% succinate (for Y. lipolytica) or 1% glycerol (for D. magnusii) as the sole source of carbon and energy.
Isolation of mitochondria. Mitochondria from Y. lipolytica and D. magnusii were isolated by the methods designed in our laboratory and described earlier in [12] and [15], respectively, with minor modifications.
Mitochondrial preparations thus obtained were fully active for at least 4 h when kept on ice.
Oxygen consumption was monitored polarographically with a closed Clark-type oxygen electrode in a 1-ml cell in medium containing 0.6 M mannitol, 2 mM Tris-phosphate buffer, pH 7.3-7.4, 20 mM pyruvate, 5 mM malate, and mitochondrial protein (0.5 mg/ml).
Mitochondrial transmembrane potential (Δψ) was measured with a Beckman Coulter DU-650 spectrophotometer (USA) at the wavelength pair (511-533 nm) with safranine O as a Δψ-related probe [16]. The incubation medium contained 0.6 M mannitol, 0.2 mM Tris-phosphate buffer, pH 7.2-7.3, 20 mM succinate, and mitochondrial protein (0.5 mg/ml).
Mitochondrial swelling was monitored with a Hitachi-557 spectrophotometer (Japan) by recording changes in apparent absorbance at 540 nm.
Mitochondrial protein was assayed by the method of Bradford [17] with BSA as standard.
RESULTS AND DISCUSSION
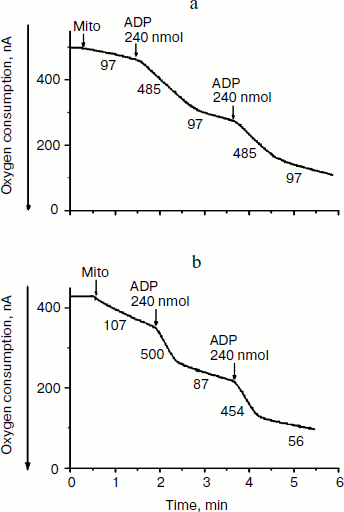
In this work we used tightly-coupled mitochondrial preparations isolated from yeasts Y. lipolytica and D. magnusii. They oxidize exogenous NAD-dependent substrates with high respiratory rates (Fig. 1), high (record) respiratory control ratios [18], and ADP/O ratios close to the theoretically expected maxima. Figure 1 depicts numerous transitions from state 4 respiration (before ADP addition) to state 3 respiration (after ADP addition), and again spontaneously to state 4 respiration after phosphorylation of the added ADP.
Respiring mitochondria maintained unchanged their Δψ for a long time (for 10-15 min) without any disturbance in light scattering even in hypotonic mannitol-based incubation medium. The absence of swelling of yeast mitochondria was not simply due to structural limitations, since large-amplitude swelling occurred in the presence of alamethicin, which being incorporated into mitochondrial membranes forms channels with a diameter of 1 nm that are permeable to divalent cations and low-molecular-weight compounds.Fig. 1. Amperometric recording of oxygen consumption by mitochondria of Y. lipolytica (a) and D. magnusii (b) respiring on pyruvate + malate. The incubation medium contained 0.6 M mannitol, 2 mM Tris-phosphate, pH 7.2-7.4, 1 mM EDTA, 20 mM pyruvate, 5 mM malate, and mitochondrial protein (0.5 mg/ml) (Mito). Respiratory control ratios upon successive additions of ADP were: a) 5.0; 5.0; b) 5.7; 8.1. ADP/O ratios upon successive additions of ADP for both mitochondrial preparations were 2.8-2.9. Numbers adjacent to traces are respiration rates in ng-atoms O/min per mg of mitochondrial protein.
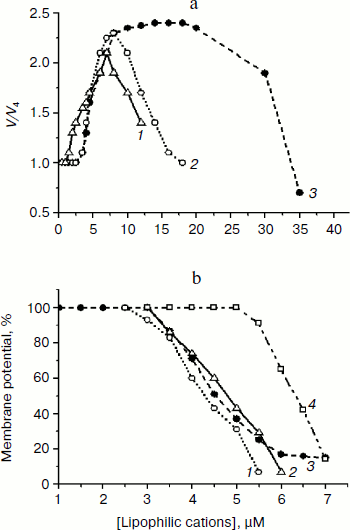
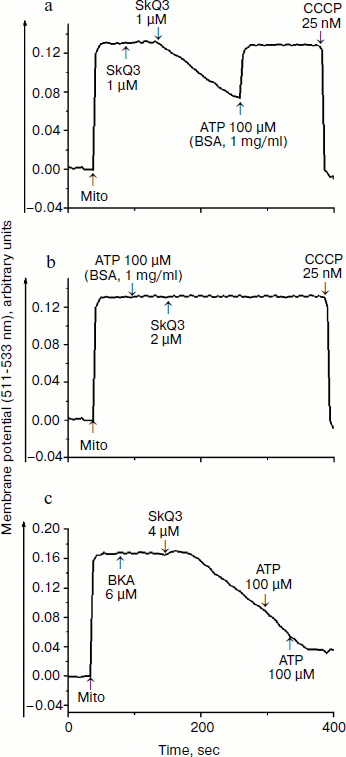
First we found that mitochondria-targeted lipophilic cations with delocalized charge (SkQ1, SkQ3, and MitoQ) activated oxidation of NAD-dependent substrates (pyruvate + malate) in state 4 respiration ~2.2 times with a half-maximal effect at 4-5 µM (Fig. 2a), which is indicative of their uncoupling effect. At higher concentrations they inhibited respiration, possibly due to their prooxidant activity. However, uncoupling observed in the presence of lipophilic cations was at least 2-3 times less effective in comparison with canonical uncouplers of the CCCP-type, suggesting that the lipophilic cations used were “relatively weak” uncouplers. Uncoupling effect of lipophilic cations was totally prevented by BSA and, to a lesser extent, by ATP (data not shown). Quite similar results were obtained upon examining the effect of lipophilic cations on the membrane potential (Fig. 2b). SkQ1, SkQ3, MitoQ, and C12TPP decreased the membrane potential generated by succinate-supported yeast mitochondria with a half-maximal effect at concentrations varying from 4 to 6.5 µM. The fact that C12TPP, the only lipophilic cation lacking plastoquinone and therefore not exhibiting prooxidant activity, decreased the membrane potential at concentrations higher than those of SkQ1, SkQ3, and MitoQ, suggests that the membrane depolarization caused by SkQ1, SkQ3, and MitoQ can be, at least partially, due to their prooxidant activity. However, the situation seems to be more complicated because C12TPP, taken at higher concentrations, did depolarize the membrane. The reduced membrane potential was almost recovered to the initial level (see, as an example, data obtained with SkQ3) by BSA and ATP (Fig. 3a). These substances were very effective in preventing uncoupling effect of lipophilic cations (Fig. 3b). “Recoupling” effect of ATP was totally blocked by atractyloside (not shown) and bongkrekic acid, inhibitors of the translocase of adenine nucleotides (Fig. 3c).
Fig. 2. Effect of mitochondria-targeted lipophilic cations MitoQ (1), SkQ1 (2), and SkQ3 (3) on respiratory rate in state 4 respiration (a) and on the membrane potential (b) generated by D. magnusii mitochondria. a) Incubation medium was as in Fig. 1. Where indicated, MitoQ (1), SkQ1 (2), and SkQ3 (3) were added. b) The incubation medium contained 0.6 M mannitol, 0.2 mM Tris-phosphate, pH 7.2-7.4, 20 mM succinate, 5 mM malate, 20 µM safranine O, mitochondrial protein (0.5 mg/ml) (Mito), and varying concentrations of SkQ1 (1), MitoQ (2), SkQ3 (3), and C12TPP (4).
Much higher concentrations of lipophilic cations were required to inhibit respiration of yeast mitochondria in state 3 (I50 values varied from 9 to 22 µM (data not shown)). The precise mechanism underlying inhibitory effect of lipophilic cations remains unknown. Most likely, it originates from their prooxidant activity. Swelling of yeast mitochondria induced by lipophilic cations, being presumably due to their prooxidant and detergent activities, was observed at concentrations exceeding 40 µM (not shown).Fig. 3. Effect of SkQ3, ATP, BSA, and bongkrekic acid (BKA) on the membrane potential generated by D. magnusii mitochondria. The incubation medium was as in Fig. 2b.
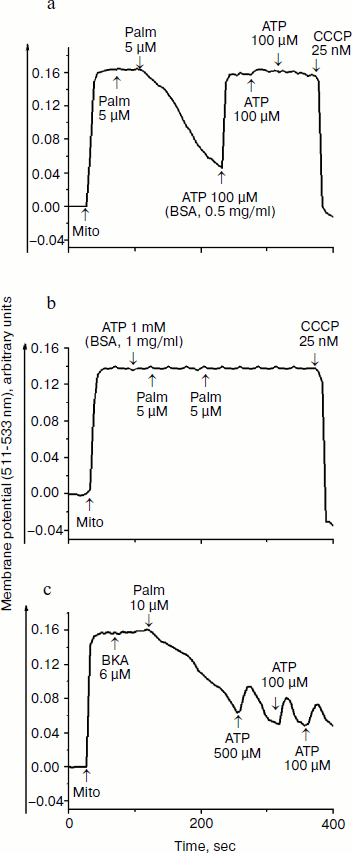
Our next step was to examine concerted action of lipophilic cations and fatty acids. Previously, we have shown [17] that both saturated and unsaturated fatty acids are poor respiratory substrates for mitochondria from Y. lipolytica and D. magnusii; in the presence of 50 µM fatty acids respiratory rates were not distinguished from those of endogenous respiration. As we noted, the specific feature of yeast mitochondria is very low, minimized rate of endogenous respiration (respiration on endogenous substrates), which allows measurement of small-scale changes in respiratory rates. In yeast mitochondria, like in animal counterparts, fatty acids act as uncouplers, as at low (micromolar) concentrations they activated state 4 respiration supported by NAD-dependent substrates (pyruvate + malate) or succinate [17]. Figure 4 depicts that fatty acids (with palmitic acid as an example) are indeed effective uncouplers as they at micromolar concentrations decreased the membrane potential, which was recovered by BSA and ATP (Fig. 4a). BSA and ATP also prevented fatty acid-induced decline of the membrane potential (Fig. 4b). Recoupling effect of ATP was almost totally blocked by atractyloside (not shown) and bongkrekic acid (Fig. 4c).
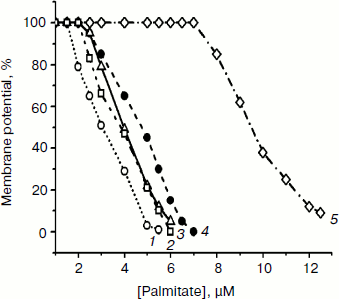
Concerted action of palmitic acid (the same results were obtained with stearic, pentadecanoic, oleic, and linoleic acids) and low concentrations of mitochondria-targeted lipophilic cations (SkQ1, SkQ3, MitoQ, and C12TPP) substantially promoted the fatty acid-induced uncoupling. In the presence of mitochondria-targeted lipophilic cations, the curve describing dependence of membrane depolarization on palmitate concentrations distinctly shifted to lower concentrations of fatty acids exerting the same uncoupling effect (Fig. 5). Inorganic phosphate, acetate, and cetyltrimethylammonium bromide (CTAB) (a lipophilic cation with localized charge) were without measurable effect (not shown). Uncoupling caused by combined action of fatty acids and lipophilic cations was partially reversed by ATP and totally by BSA (not shown).Fig. 4. Effect of palmitate (Palm), ATP, BSA, and bongkrekic acid (BKA) on the membrane potential generated by D. magnusii mitochondria. The incubation medium was as in Fig. 2b.
Thus, lipophilic cations with delocalized charges, entering mitochondria electrophoretically and accumulating within the lipid bilayer, when added to mitochondria together with fatty acids, increased proton conductance of the inner mitochondrial membrane. The most probable scenario of this relationship was put forward by V. P. Skulachev. He proposed that protonated fatty acid moves across the mitochondrial membrane by a flip-flop mechanism and then is deprotonated within mitochondria. The resulting anionic form of the fatty acid combines with a lipophilic cation, which should facilitate diffusion of the pair (fatty acid anion–lipophilic cation) across the membrane in the opposite direction. The cycle is completed by protonation of the fatty acid anion outside of mitochondria, accompanied by release of free lipophilic cation that reenters mitochondria electrophoretically. Cation accumulation within mitochondria at the expense of the membrane potential as a driving force would depolarize the membrane, which in turn should hinder the further cation accumulation, thus ensuring the mechanism of “mild” uncoupling, preventing the mitochondrial membrane potential from total collapsing.Fig. 5. Effect of varying concentrations of palmitate on the membrane potential generated by D. magnusii mitochondria in the presence of 1.5 µM SkQ1 (1), 2.5 µM C12TPP (2), 1.5 µM MitoQ (3), 1.5 µM SkQ3 (4), and when lipophilic cations were omitted (5). The incubation medium was as in Fig. 2b.
It is quite possible that the described by us and other researchers [3, 7] uncoupling induced by lipophilic cations can be explained, at least partially, by interaction of lipophilic cations with endogenous fatty acids. It is well known that mitochondria contain a pool of free fatty acids. Moreover, a positive correlation between the growth rate of an organism and the content of free fatty acids is proposed. Using the transmembrane potential as a driving force and accumulating within mitochondria, lipophilic cations can interact with endogenous fatty acids, increasing the proton conductance of mitochondria, in line with the mechanism very similar to that described for exogenous fatty acids. The essence of this mechanism is the exit from mitochondria of a pair (lipophilic cation–fatty acid), protonation of the exiting fatty acid outside of mitochondria, and release of the free lipophilic cation and its reuptake by mitochondria. The larger pool of endogenous fatty acids, the longer this cycle will be operative. In this case membrane depolarization also will assist in “self-saving” of mitochondria.
In conclusion, using as models tightly-coupled mitochondria from yeasts D. magnusii and Y. lipolytica (Figs. 2-5 present the results of examination of mitochondria from D. magnusii, but very similar results were obtained for Y. lipolytica mitochondria), we found that mitochondria-targeted lipophilic cations at low, micromolar, concentrations are “relatively weak, mild uncouplers”, at higher concentrations they inhibited respiration, at much higher concentrations they induced swelling of mitochondria, possibly due to their prooxidant and detergent actions. At very low, not uncoupling concentrations, mitochondria-targeted lipophilic cations profoundly potentiated uncoupling effect of fatty acids. We believe that these data reinforce the validity and utility of yeast mitochondria in studies of mechanisms underlying action of mitochondria-targeted lipophilic cations.
This study was supported by the Russian Academy of Sciences (program on “Molecular and Cell Biology”) and by the Russian Foundation for Basic Research (grant Nos. 06-04-49687 and 09-04-01238).
REFERENCES
1.Zorov, D. B., Isaev, N. K., Plotnikov, E. Y.,
Zorova, L. D., Stelmashook, E. V., Vasileva, A. K.,
Arkhangelskaya, A. A., and Khryapenkova, T. G. (2007) Biochemistry
(Moscow), 72, 1115-1126.
2.Harman, D. (1956) Gerontology, 11,
298-300.
3.Skulachev, V. P., Anisomov, V. N., Antonenko, Yu.
N., Bakeeva, L. E., Chernyak, B. V., Erichev, V. P., Filenko, O. F.,
Kalinina, N. I., Kapelko, V. I., Kolosova, N. G., Kopnin, B. P.,
Korshunova, G. A., Lichinitser, M. R., Obukhova, L. A., Pasyukova, E.
G., Pisarenko, O. I., Roginsky, V. A., Ruuge, E. K., Senin, I. I.,
Severina, I. I., Skulachev, M. V., Spivak, I. M., Tashlitsky, B. N.,
Tkachuk, V. A., Vyssokikh, M. Yu., Jaguzhinsky, L. S., and Zorov, D. B.
(2009) Biochim. Biophys. Acta, 1782, 437-461.
4.Murphy, M. P., and Smith, R. A. J. (2007)
Annu. Rev. Pharmacol. Toxicol., 4, 629-656.
5.Skulachev, V. P. (2007) Biochemistry
(Moscow), 72, 1385-1396.
6.Green, D. E. (1974) Biochim. Biophys. Acta,
346, 27-78.
7.Antonenko, Y. N., Avetisyan, A. V., Bakeeva, L. E.,
Chernyak, B. V., Chertkov, V. A., Domnina, L. V., Ivanova, O. Yu.,
Izyumov, D. S., Khailova, L. S., Klishin, S. S., Korshunova, G. A.,
Lyamzaev, K. G., Muntyan, M. S., Nepryakhina, O. K., Pashkovskaya, A.
A., Pletjushkina, O. Y., Pustovidko, A. V., Roginsky, V. A.,
Rokitskaya, T. I., Ruuge, E. K., Saprunova, V. B., Severina, I. I.,
Simonyan, R. A., Skulachev, I. V., Skulachev, M. V., Sumbatyan, N. V.,
Sviryaeva, I. V., Tashlitsky, V. N., Vassiliev, J. M., Vyssokikh, M.
Y., Yaguzhinsky, L. S., Zamyatnin, A. A., Jr., and Skulachev, V. P.
(2008) Biochemistry (Moscow), 73, 1273-1287.
8.Neroev, V. V., Archipova, M. M., Bakeeva, L. E.,
Fursova, A. Zh., Grigoryan, E. N., Grishanova, A. Y., Domdina, E. N.,
Ivashchenko, Zh. N., Katargina, L. A., Khoroshilova-Maslova, I. P.,
Kilina, O. V., Kolosova, N. G., Kopenkin, E. P., Korshunov, S. S.,
Kovaleva, N. A., Novikova, Y. P., Philippov, P. P., Pilipenko, D. I.,
Robustova, O. V., Saprunova, V. B., Senin, I. I., Skulachev, M. V.,
Sotnikova, L. F., Stefanova, N. A., Tikhomirova, N. K., Tsapenko, I.
V., Shchipanova, A. I., Zinovkin, R. A., and Skulachev, V. P. (2008)
Biochemistry (Moscow), 73, 1317-1328.
9.Anisimov, V. N., Bakeeva, L. E., Egormin, P. A.,
Filenko, O. F., Isakova, E. F., Manskikh, V. N., Mikhelson, V. M.,
Panteleeva, A. A., Pasyukova, E. G., Pilipenko, D. I., Piskunova, T.
S., Popovich, I. G., Roshchina, N. V., Rybina, O. Y., Saprunova, V. B.,
Samoylova, T. A., Semenchenko, A. V., Skulachev, M. V., Spivak, I. M.,
Tsybul’ko, E. A., Tyndyk, M. L., Vyssokikh, M. Y., Yurova, M. N.,
Zabezhinsky, M. A., and Skulachev, V. P. (2008) Biochemistry
(Moscow), 73, 1329-1342.
10.Bakeeva, L. E., Barskov, I. V., Egorov, M. V.,
Isaev, N. K., Kapelko, V. I., Kazachenko, A. V., Kirpatovsky, V. I.,
Kozlovsky, S. V., Lakomkin, V. L., Levina, S. B., Pisarenko, O. I.,
Plotnikov, E. Y., Saprunova, V. B., Serebryakova, L. I., Skulachev, M.
V., Stelmashook, E. V., Studneva, I. M., Tskitishvili, O. V.,
Vasilyeva, A. K., Victorov, I. V., Zorov, D. B., and Skulachev, V. P.
(2008) Biochemistry (Moscow), 73, 1288-1299.
11.Agapova, L. S., Chernyak, B. V., Domnina, L. V.,
Dugina, V. B., Efimenko, A. Y., Fetisova, E. K., Ivanova, O. Y.,
Kalinina, N. I., Khromova, N. V., Kopnin, B. P., Kopnin, P. B.,
Korotetskaya, M. V., Lichinitser, M. R., Lukashev, A. N., Pletjushkina,
O. Y., Popova, E. N., Skulachev, M. V., Shagieva, G. S., Stepanova, E.
V., Titova, E. V., Tkachuk, V. A., Vasiliev, J. M., and Skulachev, V.
P. (2008) Biochemistry (Moscow), 73,
1300-1316.
12.Kovaleva, M. V., Sukhanova,. E. I.,
Trendeleva,. T. A., Zyl’kova, M. V.,
Uralskaya,. L. A., Popova, K. M., Saris, N.-E., and
Zvyagilskaya, R. A. (2009) J. Bioenerg. Biomembr., 41,
239-249.
13.Zvyagilskaya, R., Andreishcheva, E., Soares, I.
M., Khozin, I., Berhe, A., and Persson, B. L. (2001) J. Basic
Microbiol., 41, 283-303.
14.Zvyagilskaya, R. A., Zelenshchokova, V. F.,
Ural’skaya, L. A., and Kotel’nikova, A. V. (1981)
Biokhimiya, 46, 3-10.
15.Bazhenova, E. N., Deryabina, Yu. I., Eriksson,
O., Zvyagilskaya, R. A., and Saris, N.-E. L. (1998) J. Biol.
Chem., 273, 4372-4377.
16.Akerman, K. E. O., and Wikstrom, M. K. F. (1976)
FEBS Lett., 68, 191-197.
17.Bradford, M. M. (1976) Anal. Biochem.,
72, 248-254.
18.Chance, B., and Williams, G. R. (1955)
Nature, 175, 1120-1121.