REVIEW: Classification of Phenoptotic Phenomena
G. Libertini
Independent researcher; E-mail: giacinto.libertini@tin.it
Received April 9, 2012
Phenoptosis is defined as the programmed death of an organism. In a more detailed formulation of the concept, it is the death of an individual caused by its own actions or by actions of close relatives (and not by accidents or age-independent diseases), which is determined by genes that are favored by natural selection and in certain cases increase the evolvability of organisms. This category of phenomena cannot be justified in terms of individual selection and needs always a justification in terms of supra-individual selection. Four types of phenoptosis are proposed (A, obligatory and rapid; B, obligatory and slow; C, optional; D, indirect). Examples of each type and subtype are given. The classification is discussed in its meaning and implications, and compared with another classification of end life types largely based on the classical concept of senescence.
KEY WORDS: phenoptosis, supra-individual selection, senescence, agingDOI: 10.1134/S0006297912070024
In the first Darwinian concept of natural selection, a character is favored when it determines a greater fitness for the survival or for the reproduction capacity of the individual having it [1].
In this view, after the discovery that characters are determined by genes, a general formula illustrating the frequency variation from a generation to the next, determined by natural selection, of a gene C (ΔC) acting in an individual I, could be:
where S is advantage/disadvantage for I caused by the gene C, and P is residual capacity of I of having a progeny at the age when the gene manifests its action (reproductive value). But, this formula does not explain the unselfish actions of parental care, or the cases in which reproduction entails obligatorily the death of a parent or when the adult individual decays more or less rapidly just after the reproduction. In these cases, with a somehow strained interpretation, it is necessary that the meaning of the term S is not limited to the strict individual advantage, but includes the advantages/disadvantages determined by the effects of gene C on other individuals genetically related.
A solution for these difficulties is the concept of “inclusive fitness” [2-5], where the calculation of the selective forces considers all the individuals for which the actions of a character (that is, of the gene(s) determining it) have some effect:
where n is number of individuals for which the character has some effect, Sx is advantage/disadvantage for the individual x, Px is reproductive value of individual x, and rx is coefficient of relationship between individual x and individual I.
This formula explains very well parental care and many other unselfish (and selfish) behaviors, included the cases of parental deaths connected to reproduction. The concept of inclusive fitness (kin selection) is not alternative to the classic view, but is an extension of it: individual fitness is a particular case of inclusive fitness where only individual I is involved in the effects of the character. In mathematical terms, with n = 1 and considering that r1 = 1, formula (2) coincides with formula (1).
Kin selection concepts have been largely used for some decades to explain also eusociality, in particular by using the “haplodiploid hypothesis” as justification of eusociality in many Hymenoptera species of ants, wasps, and bees. But, formulas based on the inclusive fitness concept become unworkable when multiple synergistic effects in the interactions between individuals must be considered. Moreover, many not haplodiploid species show eusociality (e.g. termites), and “the association between haplodiploidy and eusociality fell below statistical significance” [6]. It has been argued that “standard natural selection theory in the context of precise models of population structure represents a simpler and superior approach” [6].
On the other side, kin selection applied to populations composed by a single or few clones is a form of group selection that does not imply unacceptable postulates. And it is possible to propose other cases where this or that type of mathematical/logical approach could be admissible.
This small preamble does not want to discuss the validity and the limits of the various methods and concepts of population genetics used to describe and study actions and behaviors implying unselfish actions, but only to underline two common features:
– it is important to consider the possible effects of the actions of a character, or a gene, on other individuals, namely it is necessary to consider mechanisms of supra-individual selection, which can be variously defined and calculated (kin selection, precise models of population structure, group selection, mechanisms that favor the rate and the possibilities of evolution, or evolvability [7], conditions that favor semelparity or parent sacrifice, etc.), although this does not exclude that in most cases a character, or gene, has no effect on other individuals;
– the selection may favor in particular cases a character, or a gene, that is damaging for the survival of the individual where the character acts. This does not exclude that in most cases a character favored by natural selection is not damaging for individual fitness.
These general ideas are not new in the Darwinian concept of evolution, as the same Darwin said: “A tribe including many members who … were always ready to aid one another, and to sacrifice themselves for the common good, would be victorious over most other tribes; and this would be natural selection” [8].
Moreover, the idea of individual sacrifice for the common good is as old as human civilization, and it was even expressed by a philosophical mind in terms that somehow anticipate Darwinism, as already underlined in a recent meeting [9]: “Schopenhauer wrote: The individual is … not only exposed to destruction in a thousand ways from the most insignificant accidents, but is even destined for this and is led towards it by nature herself, from the moment that individual has served the maintenance of the species [10]. Today, this statement needs only one specification, i.e. the term “species” should be replaced by “species-inherent genetic program”. As a rule, interests of an individual coincide with those of the genetic program, which requires the individual to exist, multiply, and evolve. However, in certain cases, these two kinds of interests are opposite, so the genetic program forces the individual to operate in a way that is counter-productive for the individual. In extreme cases, ... it favors elimination, rather than survival, of an individual”.
DEFINITION OF “PHENOPTOSIS”
It is incredible that until few years ago, namely up to the suggestion of the neologism “phenoptosis” by Skulachev in 1997 [11, 12], in the immense scientific vocabulary there was no term indicating the death of an individual, when not determined by accidents or age-independent diseases, as an event provoked by particular mechanisms genetically regulated, namely programmed, and therefore somehow favored by natural selection. This is even stranger if we consider that, for many years prior to 1997, innumerable cases of individuals dying by the action of mechanisms clearly programmed were known in the animal and vegetable worlds. For example, in the monumental and very well documented 1990 Finch’s textbook [13], a whole long chapter is devoted to “rapid senescence and sudden death”, but the author did not consider it necessary to coin a specific name for this frequent type of events.
It is true that, relatively only a few years before, there was the description of apoptosis [14], namely the death of a cell not caused by accidents but as a phenomenon genetically programmed and having a set of functions that is adaptive. (Later, other forms of “programmed cell death” have been defined and studied, but this is a topic beyond the scope of this work.) Moreover, the description of programmed cell death in prokaryotes (“proapoptosis” [15]) and the definition of the programmed death of a mitochondrion (“mitoptosis” [16]) were even more recent.
Really, it must be considered the scientific and cultural resistance to accept fully the notion that natural selection for a character can lead to phenomena such as promoting the death of the individual having the character: seemingly, this is the exact opposite of the pivotal concept of Darwinism, namely that natural selection favors everything useful for survival and reproduction of an individual, whereas what is harmful to survival and reproduction is opposed.
However, beyond these considerations, in the original definition of Skulachev, “phenoptosis” is “the programmed death of an individual” [12]. The term was coined in analogy with apoptosis, mitoptosis, and proapoptosis. The following term “organoptosis” [17] was coined in analogy with them. A more detailed definition of the concept is as follows: “Phenoptosis is the death of an individual caused by its own actions or by actions of close relatives (siblicide; in particular, the parent-caused death of an offspring or filial infanticide) and not caused primarily by accidents or diseases or external factors, which is determined, regulated, or influenced by genes favored by natural selection”.
Phenoptosis cannot be justified in terms of individual selection and needs always a justification in terms of supra-individual selection. On the contrary, a death with no explanation in terms of supra-individual selection must have specific non-selective determinants.
As parallel considerations:
– apoptosis in unicellular organisms and proapoptosis in eubacteria, when these phenomena have an adaptive meaning, are synonyms of “phenoptosis” for the individuals killed by these phenomena;
– in all its functions in multicellular eukaryotes, apoptosis and other forms of programmed cell death can be considered as analogous phenomena or, better, an evolution of the death of a unicellular individual in a clone where the reproduction is reserved to specialized cells. Moreover, an analogy is possible with the sacrifice of an individual in a eusocial species where the reproduction is reserved to a few individuals;
– organoptosis is the organized death of many cells by apoptosis, and should be considered not a different phenomenon but only as a coordinated manifestation of apoptosis in many cells;
– on the contrary, mitoptosis is a similar but different phenomenon and should be analyzed in the awkward evolutionary context of the interactions between a complex host (the multicellular eukaryote) and clones of symbionts (the mitochondria in a cell).
CLASSIFICATION OF PHENOPTOTIC PHENOMENA
Here, a possible classification of phenoptotic phenomena is proposed:
A) obligatory and rapid phenoptosis:
A-1) related to the reproductive cycle;
A-2) deriving in general from characteristics of the life cycle;
B) obligatory and slow phenoptosis:
B-1) duplications-related increasing probability of apoptosis in unicellular eukaryotes;
B-2) age-related increasing mortality in multicellular eukaryotes;
C) optional phenoptosis:
C-1) determined by biochemical mechanisms;
C-2) determined by behavioral mechanisms;
D) indirect phenoptosis:
D-1) determined by biochemical mechanisms;
D-2) determined by behavioral mechanisms;
and, on the contrary:
E) absence of phenoptosis:
E-1) with high constant mortality rate;
E-2) with small or moderate constant mortality rate;
E-3) with age-related decreasing mortality rate.
The aim of this classification is not the definition of arbitrary boundaries between types and subtypes of the phenomenon, but to underline different types of phenoptosis that generally need different explanations in terms of natural selection. At the end of the classification of phenoptotic phenomena, the absence of them is opposed as a last different category.
A. Obligatory and rapid phenoptosis. Phenoptosis is defined as obligatory and rapid when, as a rule, it happens in all the individuals of a species in a relatively short time. This type of phenoptosis is in the range of the phenomena defined by Finch as “rapid senescence and sudden death” [13]. In some cases of semelparity and rapid senescence, the phenomenon is triggered by particular environmental or physiological signals [13], but this is only a temporal modulation of a fixed life cycle and not a form of optional phenoptosis (see below). The concept of sudden phenoptosis does not imply a short lifespan, e.g. “various species of the thick-stemmed bamboos (Phyllostachys) have prolonged phases of vegetative growth that last for many years or decades (7, 30, 60, or 120 years) according to the species, before suddenly flowering and dying ...” [13].
Subtype A-1. Related to the reproductive cycle. Examples:
– semelparity and sudden death after reproduction in many species of Salmoniformes and Anguilliformes, in some species of dasyurid marsupials and of rodents, and in many species of plants, in particular monocarpic angiosperms [13]. In particular, Finch says: “Many botanists emphasize that plant senescence is an orderly and active process (Leopold, 1961; Noodén, 1988a, 1988b, 1988c)” [13];
– endotokia matricida, that is maternal death as the obligatory result of birth, shown by some invertebrates, in which “the young kill their mother by boring through her body wall” or cannibalizing her body [13].
Subtype A-2. Deriving in general from characteristics of the life cycle. Examples:
– aphagy in adult insects: “Aphagy from defective mouthparts or digestive organs is very common during the adult phases of insects (Weismann, 1889b; Metchnikoff, 1915; Norris, 1934; Brues, 1946; Wigglesworth, 1972; Dunlap-Pianka et al., 1977) and is the limiting factor in the adult lifespan of many short-lived species. This phenomenon is, inarguably, programmed senescence ...” [13];
– lack of anatomic parts in male rotifers [13].
B. Obligatory and slow phenoptosis (or, shortly, slow phenoptosis). Phenoptosis is defined as obligatory and slow when it is characterized by an age-related progressively increasing probability of death, that is a progressively decreasing fitness. The expression “slow phenoptosis”, in relation to the age-related increasing mortality shown by many species, was proposed by Skulachev [9, 18].
Subtype B-1. Duplications-related increasing probability of apoptosis in unicellular eukaryotes. Example:
– in the mother cell lineage of yeast, the death by apoptosis follows an exponential dynamics [19] that mimics the increment of mortality rate in multicellular eukaryotes. In Finch’s classification of senescence phenomena [13] this subtype of phenoptosis is classified in the chapter “Rapid senescence and sudden death”, section “Reproduction-related rapid senescence and sudden death”, as the deaths are related to the reproduction-related duplications in the mother cell lineage.
This phenomenon has been suggested as adaptive [20]: “apoptosis coupled to chronological and replicative aging limits longevity that would maintain ancient genetic variants within the population and, therefore, favor genetic conservatism”.
Lewis argues against this interpretation [21] with the argument that a yeast cell of the mother lineage dies by apoptosis after n duplication (n = 25-35 in laboratory conditions [22]) and that the death of a single individual among 2n = 107-1010 descendants is irrelevant for any hypothesis considering the phenomenon as somehow favored by natural selection. Against this argument: it is important not the death of a single individual among innumerable descendants, but the exponentially progressive increasing probability of apoptosis, which causes a quicker generation turnover and contrasts the “genetic conservatism” mentioned by Büttner et al.
Subtype B-2. Age-related increasing mortality in multicellular eukaryotes. Examples:
– many species of multicellular eukaryotes show an “increasing mortality with increasing chronological age in the wild” [23, 24], defined with its acronym (IMICAW [25]) or as “actuarial senescence” [26] or as “age-related fitness decline in the wild” [27], or described as “gradual senescence with definite lifespan” [13].
The first three definitions are descriptive and imply no explanation for the phenomenon. On the contrary, according to the common meaning attributed to the term senescence/aging, namely the unavoidable age-related deterioration of everything, both animate or inanimate, Finch’s definition seems to mean, intentionally or not, a gradual unavoidable deterioration caused by factors not determined by natural selection. On the other side, the definition “slow phenoptosis” used in this classification assumes a selective advantage for the phenomenon, as underlined in the section “Definition of Phenoptosis”.
There are some facts:
–age-related progressive fitness decline is well documented in natural conditions. On the basis of Ricklefs’ data [28] it is possible to define the life tables for many animals (see figure). As this phenomenon exists in natural conditions, it is subject to natural selection and, therefore, needs an explanation based on the selective pressures – positive and negative, individual and supra-individual – to which it is subject;
– in the wild, the older individuals, which show the advanced signs of what is commonly said senescence, are rare or, in their more advanced expressions, not-existent; but this fact does not contradict the existence in the wild of age-related mortality increase;
– about the phenomenon, it has been stated that it cannot be explained by selection as it is certainly harmful to survival and reproductive capacity of the individual showing it [29]. This consideration is unacceptable as it disregards possible explanations based on supra-individual types of selection.
C. Optional phenoptosis. Phenoptosis is defined as optional when it is triggered only in particular conditions on the basis of genetically determined mechanisms that favor or oblige the phenomenon.Life tables and death rates in wild conditions of hippopotamus Hippopotamus amphibius (a) and lion Panthera leo (b) (data from Ricklefs [28])
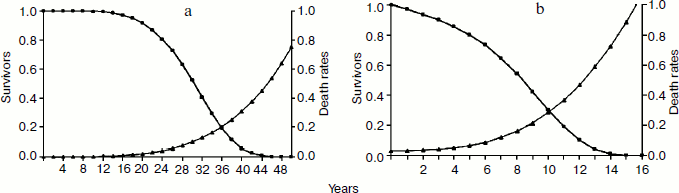
Subtype C-1. Determined by biochemical mechanisms. Phenoptosis is triggered only in particular conditions in which it entails an advantage for individuals of the same group/deme/tribe (likely related) or of the same clone (by definition related). Examples:
Proapoptosis in eubacteria as:
– bacterial phytoplankton mass suicide as defense against viruses [30]. In particular, “as most plankton in a bloom are near identical genetically, from the perspective of their genes, a die-off that creates enough scorched earth to stop the viral advance can make sense” [30];
– bacterial suicide triggered by phage infection, “thereby curtailing viral multiplication and protecting nearby E. coli from infection” [31];
– the “built-in suicide module” activated by antibiotics in E. coli [32] and other bacteria [21].
The programmed cell death in prokaryotes has been defined as “proapoptosis” and hypothesized as phylogenetic precursor of eukaryotic apoptosis [15], with which it shares various features [33]. The fact that this type of phenomena is genetically determined or programmed has been emphasized [17, 21].
Apoptosis in unicellular eukaryotes. In yeast, an individual, composed by a single cell, divides in a mother cell and a daughter cell. For the mother cell lineage, in relation to the number of duplications there is an increasing vulnerability to apoptosis [20, 34-36], and the death rate increment follows exponential dynamics [19]. In combination with this increasing propensity to apoptosis, it is triggered by: a) unsuccessful mating [20]; b) dwindling nutrients [37]; c) chemical alterations [38].
Cellular fragments of individuals that have died by apoptosis do not damage nearby cells and are usefully phagocytized by them, which, therefore, “are able to survive longer with substances released by dying cells” [35].
Yeast apoptosis is interpreted as adaptive, being useful to the survival of the clone, likely made up of kin individuals [16, 17, 35, 39-43]. On the contrary, the apoptosis triggered by toxin secreted by competing yeast tribes [20] is not adaptive for the attacked tribe and clearly means an exploitation of apoptotic mechanisms by the assailants.
Subtype C-2. Determined by behavioral mechanisms. In particular conditions, selection favors behaviors that are risky or deadly for the individual showing it, but that increase survival probability of related individuals. The adjective “behavioral” means that the mechanisms require the action of a nervous system, with or without the action of instinct and/or intelligence and/or awareness, however defined or conceived. The adjective does not imply that the functions of a nervous system are not based on or influenced by biochemical mechanisms, hormones included. Examples:
Unselfish and deadly behaviors in invertebrates. Behaviors of individual sacrifices in eusocial insect species (ants, bees, termites, etc.) are well-known [5].
Unselfish and deadly behaviors in vertebrates. Unselfish behaviors that jeopardize one’s survival for the benefit of others are common in social vertebrate species. For example, with great individual risk, the predominant males of yellow baboons (Papio cynocephalus) [44] and chacma baboons (Papio ursinus) [45] place themselves in the most exposed positions to defend their drove from predators.
For various species of birds, there are distraction behaviors shown by parents with great individual risk to save offspring threatened by predators [46-48].
Unselfish and potentially deadly behaviors in man. There are countless cases in which one or more individuals sacrifice their own life, or at least endanger it seriously, to save the lives of others. When a man does such acts of sacrifice, this is usually attributed to the choice of the individual, as an expression of free will, and not to mechanisms determined by genes because this second interpretation would be in fact a denial of the free will. The question becomes religious and philosophical and any scientific value is lost. However, a plausible scientific thesis is that, both for eusocial vertebrates and for man, there are not genes that determine rigidly particular behaviors under specific conditions, but that particular combinations of genes determine a neuroendocrine development (or whatever it is correct to describe it) that under certain conditions tends to favor certain types of reaction or strategies.
For example, in a state of imminent danger, some will die trying to save those around them, and others will flee in the attempt to save their own lives, even if this affects the survival chances of others. The choice between these two opposing strategies is also strongly influenced by the degree of relationship between the individual who must choose the strategy and the people whose survival is threatened: it is very likely that the first strategy is chosen if their offspring are threatened and less likely to do so if the contrary is true.
In animals, analogous behaviors are interpreted as determined by instincts, namely it is recognized that in some way the behavior has been shaped by natural selection. In humans, while acknowledging that in large part they are determined by acts of will, it would be objective to admit that there are instinctive components determined by natural selection as well as for the animals. Moreover, wanting to be stringent, for the part that is determined by the will, perhaps it is not wrong to argue that intelligence, consciousness, will, and all those characters that somehow make us different from other evolutionarily near species, are a result of natural selection and therefore, although more indirectly, were shaped by similar selection pressures.
D. Indirect phenoptosis. Phenoptosis is indirect when, in particular conditions, there is the death of an individual caused by its close relative, in particular the case of the parent-caused death of an offspring.
Subtype D-1. Determined by biochemical mechanisms. Examples:
– in the mouse, a new partner of a female kills the new-born offspring. So, when a new male takes over, a female aborts its own young. This is interpreted as adaptive because it saves for the mother time and energy, deriving from the predictable killing of the young after birth [49];
– in vertebrates, it is indispensable that the immune system discriminates between antigens of each host individual and those of the parasites, which try to overcome immunologic defenses by using for their coverings proteins with the same antigenicity of the host (antigen mimicry). The defense of the host against antigen mimicry is to have the greatest inter-individual variability of antigen formulas so that a mimicry adapted to infect all the potential hosts is impossible. The major histocompatibility complex (MHC) is the main tool by which the host organism obtains extraordinary antigen variability. Differences between antigenic formulas of host and parasite give greater resistance to infection, while similarities cause susceptibility. Correlations between resistance or susceptibility to several infectious or infection-related diseases and specific human MHC alleles are well documented [50, 51].
As the best progeny is that with the greater antigen variability, MHC-mediated mate choice and post-copulatory selection try to achieve this result. The first case is widespread in nature and is documented for several vertebrate taxa [52], and also for our species: a) women college students rated the odors of MHC-dissimilar men as being “more pleasant” than those of MHC-similar men [53, 54]; b) in an isolate, ethnically homogenous community, significantly fewer couples were observed to match at a 16-locus MHC haplotype [55, 56].
The second case, also defined as “cryptic female choice” [57], spontaneous and non-pathologic miscarriages eliminate the offspring with lesser antigen variability with a decreased fitness due to reduced potential resistance to infective diseases [58]. Post-copulatory selection is well documented in animals [59]. A study on human subjects documented an excess of MHC-heterozygotes in newborn males [60]. Several studies on an ethnically homogenous and isolated community documented that couples with shared HLA-DR alleles in comparison with couples not sharing the same alleles have: 1) significantly fewer children [61]; 2) a greater interval between pregnancies [62]; 3) a greater pregnancy loss rate [63];
– “... sonograms of women in the first trimester of pregnancy reveal that twins are conceived two to four times more often than they are born; in the majority of cases, the smaller of the two fetuses disappears by the third trimester and is apparently reabsorbed by the mother (Robinson and Caines, 1977; Varma, 1979)” [64]. The phenomenon (the “vanishing twin”, reviewed in 1998 [65]) should be evaluated considering that one of the common determinants of filial infanticide (see later) is the twin birth as scant resources do not allow the successful breeding of two children at the same time.
Subtype D-2. Determined by behavioral mechanisms. For the meaning attributed to the adjective “behavioral”, see subtype C-2. Filial infanticide is the killing of an offspring by its own parents. Examples:
– for our species, the abandonment of healthy new-born babies or direct filial infanticide when the resources are insufficient are widespread and ancient behaviors [66]. The plausible evolutionary interpretation of these acts, which are present even in modern societies, is that progeny with reduced survival possibility subtracts precious resources to parents and kin individuals [67];
– in the animals, filial infanticide is widespread and often involves cannibalism [64].
E. Absence of phenoptosis. Slow phenoptosis is absent when in the wild the mortality rate does not increase in relation with age1.
1“However, the presence of an age-dependent mortality increase does not obligatorily mean that such an effect is a result of phenoptosis. For example, in the lens proteins (crystallins) of big whales spontaneous isomerization of L-amino acids to D-amino acids occurs at any age. Crystallins are synthesized during formation of the lens and originally contain, as any other proteins, only L-amino acids. Crystallins are practically not replaced during the entire life of the whale. L-D isomerization is not encoded by genomes and it is a chemical property of amino acids. Fortunately, this process is very slow (2% per 10 years). However, after 200 years, 40% of L-amino acids are already isomerized to D-amino acids in crystallins, an event strongly affecting the spatial structure of these proteins and apparently their unique ability to be absolutely transparent for the visible light. If such a process results in formation of cataract, it may lead to blindness, which should make impossible the life of an old whale in the ocean. He should die, such a death being age-dependent. And nevertheless, it cannot be regarded as slow phenoptosis” [68]. (Comment of the Editor-in-Chief)
Subtype E-1. With high constant mortality rate. In this case, which is very frequent among the insects and many other invertebrates as the famous C. elegans, in the wild the mortality is so high that it is rare or virtually not-existent the possibility that an individual reaches the ages at which, in protected laboratory conditions, it is possible to observe a progressive increase of mortality [13]. This “increasing mortality with increasing chronological age in captivity” (IMICAC [25]) is clearly different from the IMICAW phenomenon because, being absent in natural conditions, it cannot be influenced by natural selection, while the contrary is true for IMICAW. However, as for these species generally there is almost neither cell turnover nor capacity of repairing parts of their soma that are worn out or damaged, it could be discussed if the characteristics of their life cycle should be classified within the category A. Obligatory and rapid phenoptosis, subtype A-2. Deriving in general from characteristics of the life cycle. In the textbook [13], Finch chose to classify these phenomena within the category “rapid senescence and sudden death”, but it is necessary to emphasize that many of the observations reported by him are referred to ages existing only in laboratory conditions.
Wanting an exact observance of the classification criteria of this work, which refers to phenoptosis phenomenon and not to senescence in its broad meaning as in the case of Finch’s textbook, if for a species the death deriving from an absent cell turnover or from unrepaired mechanical wear or damage happens in natural conditions (at least in a non-minimal percentage), we should choose the classification in the subtype A-2, because in this case it is a programmed part of the life cycle, as it is directly influenced by natural selection. On the contrary, if the death for the above-said causes is practically a laboratory phenomenon, the classification should be in the present subtype E-1, because the death is due to the absence of a selection useful for a life extended to ages non-existing in the wild. However, the difference is unimportant in its essence. In both interpretations, the natural selection cannot or does not act for a greater duration of the life, e.g. favoring mechanisms of cell turnover or of repair of parts worn out or damaged, because the life cycle is such that a greater duration of life does not involve any advantage.
Subtype E-2. With small or moderate constant mortality rate. Finch provides a broad overview of animal and plant species for which in natural conditions an age-related fitness decline, alias increased mortality, is not observed and this condition is defined as “negligible senescence” [13], that is an absence of signs of aging to an extent to be detectable or statistically significant. The absence of an age-related increasing mortality for a species does not mean that the individuals of the species are immortal. After a certain time T, in function of the mortality rate, the probability that an individual is still alive is minimal. Besides, from that age T, by definition natural selection cannot have an impact, and it is impossible that a harmful event acting after age T can favor the development of a character contrasting the damage.
For the supporters of the idea that aging is something inevitable, this is interpreted as a confirmation of the idea. For the supporters of the contrary idea that aging is something genetically programmed, the phenomenon of functions decay for species not showing such a decline at ages existing in the wild and then showing the decline at later ages existing only in captivity conditions, is indicated with a specific name (IMICAC) and is not confused with the phenomenon IMICAW [25].
Subtype E-3. With age-related decreasing mortality rate. The phenomenon for which the mortality rate decreases in relation with the age, also defined “negative senescence” [69], is shown by some species for which there is no function decline and, on the contrary, other factors, as a greater soma, causes a reduced predation and, so, an age-related decreasing mortality.
The definition “negative senescence” is misleading, as one could understand it as a form of reverse aging. In fact, it is a case of small or moderate constant mortality rate (subtype E-2) with the addition of a reduction of mortality due to other factors, as a reduced predation. Therefore, subtype E-3 should be considered only a variant of the previous subtype.
COEXISTENCE OF SEVERAL TYPES OF PHENOPTOSIS IN THE SAME
SPECIES
A single individual can die only once, but the many individuals of a species can die according to more than one type of phenoptosis as there is no reason for which several types of phenoptosis cannot coexist in a single species.
In particular, limiting as an example the discussion to our species, we are subject to: optional phenoptosis (subtype C-2: Determined by behavioral mechanisms); slow phenoptosis (subtype B-2: Age-related increasing mortality in multicellular eukaryotes); indirect phenoptosis (subtype D-1: Determined by biochemical mechanisms, and D-2: Determined by behavioral mechanisms).
Moreover, within our body there are numberless cases of phenomena that are similar to phenoptosis: a) every day, countless apoptotic events for cells in turnover; b) in particular cases, mitoptosis; c) in morphogenetic phases, organoptosis; and d) very likely, within the billions of eubacteria living on our teguments and inside our cavities, cases of proapoptosis.
GENERAL SCHEMA FOR THE STUDY OF A PHENOPTOTIC PHENOMENON
First of all, it is necessary to establish that a phenomenon X is within the limits of phenoptosis definition.
Then, it is important to distinguish between:
– primary causes, that is the evolutionary determinants and the specific genetic mechanisms favored by natural selection;
– secondary causes, that is the physiological mechanisms that determine and modulate the phenomenon;
– tertiary causes, that is the final causes of death.
For example, in aphagous insects, which in the adult stage lack mouthparts or digestive organs and can survive only a limited time, we have: i) evolutionary determinants that favor such a strange (for us vertebrates) condition; ii) molecular mechanisms, genetically determined and regulated, which cause the above said defects; iii) the consequent deadly starvation.
Finally, it is necessary to compare similar species where phenoptosis is present only in some of them (or, for a single species, only in particular conditions), and study the evolutionary (primary causes) and physiological (secondary causes) determinants of the presence/absence of the phenomenon.
For example, in Salmoniformes, 10 out of 90 genera show semelparity followed by death while the other genera show iteroparity [13]; Alosa sapidissima, a herring, show semelparity at lower latitudes “while iteroparity increases linearly with the degree latitude” [13].
The schematic classification of the phenoptotic phenomena exposed in this paper is not at all in competition for completeness and documentation with Finch’s textbook that covers broadly the endless variety of ways according to which individuals of different species end their lives [13] or with analogous manuals.
The approach of the two types of classification is significantly different and must be somehow discussed.
Finch describes the modes of the end of life, defining them as different types of senescence. It is implicit in this approach the concept that all bodies undergo senescence: the fact that many species of animals and plants do not show detectable signs of aging is exposed by Finch, but no plausible and/or general explanation is given. On the contrary, the classification exposed in this paper is based on the concept that each phenoptotic phenomenon shown by a species presumes a definite evolutionary advantage, because without it the phenomenon could not exist and, so, the general principle, according to which natural selection favors those who best survive, should be applied.
In this alternative view, the default condition favored by natural selection is the absence of age-related fitness decline, obviously in ages present in wild conditions, namely what Finch calls “negligible senescence” but that should be called “absence of senescence”: on the contrary, the presence of phenomena for which the fitness is reduced, suddenly or slowly, always or in particular conditions, is necessarily due to special evolutionary necessities that somehow justify their existence.
It should be noted that, in most cases of the phenoptotic phenomena, the prevailing view in the scientific world is that they are determined by specific evolutionary necessities. The only big exception to this common vision is the slow phenoptosis for multicellular eukaryotes, namely what is commonly called aging. For this particular category of events, the prevailing opinion is that they are the result of insufficient selection for a longer lifespan.
A few isolated heretics, a tiny minority, argue with various arguments and in various ways that slow phenoptosis is the active result of natural selection, as it is for other types of phenoptosis, and not the outcome of insufficient selection for a longer lifespan.
It is ironic to note that these isolated heretics in the wider context of the scientific opinions about phenoptotic phenomena are part of a large majority and, on the contrary, the non-heretics about the slow phenoptosis are heretics in the broader context of phenoptosis.
However, the phenoptosis concept has also a great practical meaning. It is a turning point from a paradigm in which senescence is considered a non-physiological phenomenon, a sum of many different forms of decay and wear, in short a simple name over a mass of various out of control processes, to a new paradigm where aging is determined and regulated by genes, has an evolutionary meaning, physiological mechanisms, and phylogenetic correlations and is a particular expression of a broader category of phenomena, phenoptosis, which has a central importance in biology and deserves the best studies.
REFERENCES
1.Darwin, C. R. (1859) On the Origin of Species by
Means of Natural Selection, or the Preservation 1 of the Favoured Races
in the Struggle for Life, John Murray, London.
2.Hamilton, W. D. (1964) J. Theor. Biol.,
7, 1-52.
3.Hamilton, W. D. (1970) Nature, 228,
1218-1220.
4.Trivers, R. L. (1971) Quart. Rev. Biol.,
46, 35-57.
5.Wilson, E. O. (1975) Sociobiology, The New
Synthesis, Harvard University Press, Cambridge.
6.Nowak, A. M., Tarnita, C. E., and Wilson, E. O.
(2010) Nature, 466, 1057-1062.
7.Kirschner, M., and Gerhart, J. (1998) Proc.
Natl. Acad. Sci. USA, 95, 8420-8427.
8.Darwin, C. R. (1871) The Descent of Man,
John Murray, London.
9.Skulachev, V. P. (2010) The Talk at the
“From Homo sapiens to Homo sapiens liberatus” workshop,
2010, May 26, Moscow.
10.Schopenhauer, A. (1819) Die Welt als Wille und
Vorstellung: Vier Bücher, nebst einem Anhange, der die Kritik der
Kantischen Philosophie enthält, Brockhaus, Leipzig (in
English: The World as Will and Representation, Dover
Publications, New York, 1969).
11.Skulachev, V. P. (1997) Biochemistry
(Moscow), 62, 1191-1195.
12.Skulachev, V. P. (1999) Biochemistry
(Moscow), 64, 1418-1426.
13.Finch, C. E. (1990) Longevity, Senescence, and
the Genome, The University of Chicago Press, Chicago-London.
14.Kerr, J. F. R., Wyllie, A. H., and Currie, A. R.
(1972) Br. J. Cancer, 26, 239-257.
15.Hochman, A. (1997) Crit. Rev. Microbiol.,
23, 207-214.
16.Skulachev, V. P. (1999) Mol Aspects Med.,
20, 139-184.
17.Skulachev, V. P. (2003) in Topics in Current
Genetics, Vol. 3 (Nyström, T., and Osiewacz, H. D., eds.)
Model Systems in Aging, Springer-Verlag, Berlin.
18.Skulachev, V. P. (2002) Ann. N. Y. Acad.
Sci., 959, 214-237.
19.Laun, P., Bruschi, C. V., Dickinson, J. R.,
Rinnerthaler, M., Heeren, G., Schwimbersky, R., Rid, R., and
Breitenbach, M. (2007) Nucleic Acids Res., 35,
7514-7526.
20.Büttner, S., Eisenberg, T., Herker, E.,
Carmona-Gutierrez, D., Kroemer, G., and Madeo, F. (2006) J. Cell
Biol., 175, 521-525.
21.Lewis, K. (2000) Microbiol. Mol. Biol.
Rev., 64, 503-514.
22.Jazwinski, S. M. (1993) Genetica,
91, 35-51.
23.Libertini, G. (2006) The Scientific World
J., 6, 1086-1108.
24.Libertini, G. (2008) The Scientific World
J., 8, 183-193.
25.Libertini, G. (1988) J. Theor. Biol.,
132, 145-162.
26.Holmes, D. J., and Austad, S. N. (1995) J.
Gerontol. A Biol. Sci., 50, B59-66.
27.Libertini, G. (2009) in Telomeres: Function,
Shortening and Lengthening (Mancini, L., ed.) Nova Science
Publishers Inc., New York.
28.Ricklefs, R. E. (1998) Am. Nat.,
152, 24-44.
29.Kirkwood, T. B. L., and Austad, S. N. (2000)
Nature, 408, 233-238.
30.Lane, N. (2008) Nature, 453,
583-585.
31.Raff, M. C. (1998) Nature, 396,
119-122.
32.Engelberg-Kulka, H., Sat, B., Reches, M., Amitai,
S., and Hazan, R. (2004) Trends Microbiol., 12,
66-71.
33.Koonin, E. V., and Aravind, L. (2002) Cell
Death Differ., 9, 394-404.
34.Fabrizio, P., and Longo, V. D. (2008) Biochim.
Biophys. Acta, 1783, 1280-1285.
35.Herker, E., Jungwirth, H., Lehmann, K. A.,
Maldener, C., Fröhlich, K. U., Wissing, S., Büttner, S.,
Fehr, M., Sigrist, S., and Madeo, F. (2004) J. Cell Biol.,
164, 501-507.
36.Laun, P., Pichova, A., Madeo, F., Fuchs, J.,
Ellinger, A., Kohlwein, S., Dawes, I., Fröhlich, K.-U., and
Breitenbach, M. (2001) Mol. Microbiol., 39,
1166-1173.
37.Granot, D., Levine, A., and Dor-Hefetz, E. (2003)
FEMS Yeast Res., 4, 7-13.
38.Madeo, F., Fröhlich, E., Ligr, M., Grey, M.,
Sigrist, S. J., Wolf, D. H., and Fröhlich, K. U. (1999) J. Cell
Biol., 145, 757-767.
39.Fabrizio, P., Battistella, L., Vardavas, R.,
Gattazzo, C., Liou, L. L., Diaspro, A., Dossen, J. W., Gralla, E. B.,
and Longo, V. D. (2004) J. Cell Biol., 166,
1055-1067.
40.Longo, V. D., Mitteldorf, J., and Skulachev, V.
P. (2005) Nat. Rev. Genet., 6, 866-872.
41.Mitteldorf, J. (2006) Rejuvenation Res.,
9, 346-350.
42.Skulachev, V. P. (2002) FEBS Lett.,
528, 23-26.
43.Skulachev, V. P., and Longo, V. D. (2005) Ann.
N. Y. Acad. Sci., 1057, 145-164.
44.Altmann, S. A., and Altmann, J. (1970) Baboon
Ecology: African Field Research, The University of Chicago Press,
Chicago.
45.Hall, K. R. (1960) Behaviour, 16,
261-294.
46.Armstrong, E. A. (1947) Bird Display and
Behaviour: an Introduction to the Study of Bird Psychology, Lindsay
Drummond, London (reprinted by Dover, New York, 1965).
47.Brown, R. G. (1962) Ibis, 104,
1-12.
48.Gramza, A. F. (1967) Auk., 84,
72-86.
49.Bruce, H. M. (1959) Nature, 184,
105.
50.Lechler, R., and Warrens, A. (2000) HLA in
Health and Disease, Academic Press, San Diego, California.
51.Shiina, T., Inoko, H., and Kulski, J. K. (2004)
Tissue Antigens, 64, 631-649.
52.Slev, P. R., Nelson, A. C., and Potts, W. K.
(2006) Curr. Opin. Immunol., 18, 608-616.
53.Wedekind, C., Seebeck, T., Bettens, F., and
Paepke, A. J. (1995) Proc. Biol. Sci., 260, 245-249.
54.Wedekind, C., and Füri, S. (1997) Proc.
Biol. Sci., 264, 1471-1479.
55.Ober, C., Weitkamp, L. R., Cox, N., Dytch, H.,
Kostyu, D., and Elias, S. (1997) Am. J. Hum. Genet., 61,
497-504.
56.Ober, C., Weitkamp, L., and Cox, N. (1999) in
Advances in Chemical Signals in Vertebrates (Johnston, R. E.,
Muller-Schwarze, D., and Sorenson, P. W., eds.) Kluwer Academic Press,
New York.
57.Loisel, D. A., Alberts, S. C., and Ober, C.
(2008) in Evolution in Health and Disease (2nd Edn.) (Stearns,
S. C., and Koella, J. C., eds.) Oxford University Press, Oxford.
58.Apanius, V., Penn, D., Slev, P. R., Ruff, L. R.,
and Potts, W. K. (1997) Crit. Rev. Immunol., 17,
179-224.
59.Tregenza, T., and Wedell, N. (2000) Mol.
Ecol., 9, 1013-1027.
60.Dorak, M. T., Lawson, T., Machulla, H. K., Mills,
K. I., and Burnett, A. K. (2002) Genes Immun., 3,
263-269.
61.Ober, C., and van der Ven, K. (1997) in
Current Topics in Microbiology and Immunology (Olding, L. B.,
ed.) Springer-Verlag, Berlin.
62.Ober, C. (1992) Exp. Clin. Immunogenet.,
9, 1-14.
63.Ober, C., Hyslop, T., Elias, S., Weitkamp, L. R.,
and Hauck, W. W. (1998) Hum. Reprod., 13, 33-38.
64.Hausfater, G., and Hrdy, S. B. (1984)
Infanticide: Comparative and Evolutionary Perspectives, Aldine,
New York.
65.Landy, H. J., and Keith, L. G. (1998) Hum.
Reprod. Update, 4, 177-183.
66.Scrimshaw, S. C. M. (1984) in Infanticide:
Comparative and Evolutionary Perspectives (Hausfater, G., and Hrdy,
S. B., eds.) Aldine, New York.
67.Eaton, S. B., Shostak, M., and Konner, M. (1988)
The Paleolithic Prescription: a Program of Diet and Exercise and a
Design for Living, Harper & Row, New York.
68.George, J., Bada, J., Zeh, J., Scott, L., Brown,
S., O’Hara, T., and Suydam, R. (1999) Can. J. Zool.,
77, 571-580.
69.Vaupel, J. W., Baudisch, A., Dölling, M.,
Roach, D. A., and Gampe, J. (2004) Theor. Popul. Biol.,
65, 339-351.