Inhibitory Effect of Post-micellar SDS Concentration on Thermal Aggregation and Activity of Papain
A. Qadeer, M. Zaman, and R. H. Khan*
Interdisciplinary Biotechnology Unit, Aligarh Muslim University, Aligarh-202002, India; fax: + 91-571-2721776; E-mail: rizwanhkhan@hotmail.com; rizwanhkhan1@gmail.com* To whom correspondence should be addressed.
Received February 21, 2014
Papain, a cysteine protease isolated from the latex of Carica papaya, is known to undergo irreversible thermal unfolding. In this study, we found that thermal unfolding of papain is accompanied by a simultaneous self-assembly process where this protein is observed to aggregate above 50°C. The extent of aggregation increased with increasing protein concentration from 3-40 µM. The aggregation was confirmed by enhanced turbidity, light scattering intensity, 1-anilino-8-naphthalene sulfonate (ANS) fluorescence intensity and by transmission electron microscopy. Furthermore, we noted that post-micellar concentration of sodium dodecyl sulfate (SDS) remarkably suppresses the thermal aggregation of papain. Far-UV circular dichroism studies revealed that SDS significantly enhances α-helical content of the protein and also tends to prevent its unfolding, and thus inhibits aggregation. Additionally, papain showed maximal activity at 65°C in neutral buffer. However, in the presence of 6 mM SDS (above its critical micellar concentration), the enzyme lost activity by about 10-fold. Thus, promoting the helical propensity of the protein does not appear to be a suitable strategy to overcome the aggregation related problems of industrially important proteins such as papain, which are not only required to be protected against aggregation but also need to remain functionally active in the presence of aggregation inhibitors.
KEY WORDS: papain, thermal aggregation, sodium dodecyl sulfate (SDS), critical micellar concentration (CMC), circular dichroism, 1-anilino-8-naphthalene sulfonate (ANS)DOI: 10.1134/S0006297914080069
The aggregation of proteins during their purification, processing, and storage is one of the major hurdles in the large-scale production of protein-based products [1]. In addition, aggregation remains an important factor behind thermoinactivation of proteins [2], which is especially undesirable for industrially important proteins. Several environmental factors are known to influence the aggregation propensity of a protein, namely temperature, protein concentration, pH, and the presence of cosolvents and contact surfaces, to name a few [3]. Among these, temperature is probably the most common factor affecting protein aggregation [4-7]. This is because unfolding of a protein above a certain temperature leads to the exposure of hydrophobic residues, which promotes immediate aggregation [8]. Indeed, recent studies have concluded that the first step of aggregation encompasses a partial unfolding of the native conformation of a protein. Consequently, some specific regions such as hydrophobic sites or free -SH groups become more exposed to new intermolecular interactions and so contribute to the formation of aggregates [9, 10]. At present, there is no general solution to the problem of protein aggregation. However, it is well recognized that intrachain interactions facilitate specific folding of a polypeptide towards native conformation, while interchain interactions lead to protein aggregation. Therefore, promoting factors that favor the kinetic competition toward intrachain interactions may hold the key to counter this devastating process and also generate the proteins in its native state [11]. In this context, several strategies have been proposed to prevent aggregation of proteins. One such approach is the use of additives such as sugars, polyols, amino acids, amines, salts, polymers, and surfactants in the solution [12].
Ionic surfactant, particularly sodium dodecyl sulfate (SDS), is known to significantly influence the aggregation behavior of proteins depending upon its concentration. Sub-micellar concentration of SDS is found to induce strong aggregation of proteins. However, higher concentration of SDS, especially above the critical micellar concentration (CMC), is known to remarkably suppress the self-assembly process [13-15]. It is believed that aggregation inhibition at post-micellar SDS concentrations occurs mainly due to the increased helical propensity of the polypeptide chain. This leads to the proposal that promoting the formation of helical structures in proteins is likely to inhibit their aggregation as it will enhance the population of species that undergo intrachain interactions and, hence, will reduce the propensity for inter-polypeptide association [13, 16]. This strategy has been proposed mainly in the context of those proteins that undergo amyloid fibril formation that is associated with neurodegenerative diseases [14, 16, 17]. However, it has not been discussed whether this approach can be applied in the case of industrially important proteins, which are not only required to be protected against aggregation but also need to remain functionally active in the presence of aggregation inhibitors.
Papain is a model cysteine protease isolated from the latex of Carica papaya. It is an α + β class protein containing two distinct domains that are known to fold independently [18]. Papain is well studied because of its molecular structure and tremendous industrial applications including beverages production, baking processes, leather processing, silk processing in the textile industry, cosmetic preparations, chitosan processing, processing of photographic films, synthesis of detergents, and many other uses [19].
The thermal denaturation of papain as studied by differential scanning calorimetry reveals that the protein denatures irreversibly [20]. Whether or not papain undergoes aggregation during thermal denaturation is not yet reported. In this study, we used several spectroscopic techniques together with microscopic analysis and demonstrated that thermal unfolding of papain at neutral pH is also accompanied by aggregation, the extent of which increases with increase in protein concentration. Further, we observed the effect of post-micellar SDS concentration on thermal aggregation of papain and found that preincubation with SDS significantly reduces the aggregation propensity of the protein. Also, the helical structure of the protein was significantly enhanced in the presence of SDS. However, it was found that suppression of aggregation using SDS does not restore the activity of the protein. The results suggest that promoting helical structure of a protein for inhibition of aggregation may not be a useful strategy to prevent thermal aggregation of industrially important proteins/enzymes.
MATERIALS AND METHODS
Materials. Papain from C. papaya (P4762), SDS, L-cysteine, and ANS (1-anilino-8-naphthalene sulfonate) were purchased from Sigma-Aldrich (USA). All other reagents used were of analytical grade.
Sample preparation. Papain was dissolved in 20 mM sodium phosphate buffer, pH 7.0, containing 5 mM sodium tetrathionate for inactivation of the proteolytic activity. The protein solution was dialyzed extensively. Protein concentration was determined using ε1%280 nm = 25 on a Perkin Elmer Lambda 25 UV-Visible spectrophotometer [21]. The molecular weight of the protein was taken as 23,406 Da [22]. A stock solution of SDS was prepared in 20 mM sodium phosphate buffer, pH 7.0. The stock solutions and buffers were filtered through a 0.45 μm syringe filter.
pH determination. pH was measured using a Mettler Toledo pH meter (Seven Easy S20-K) using an Expert “Pro3 in 1” type electrode. The least count of the pH meter was 0.01 pH unit.
Circular dichroism. Circular dichroism (CD) was measured with a JASCO spectropolarimeter (J-815) in a cuvette of 1-mm pathlength. The instrument was calibrated with D-10-camphorsulfonic acid. The thermal unfolding of papain at different conditions was evaluated by measuring the temperature-dependent CD response at 222 nm from 20 to 90°C at the rate of 1°C/min. The far-UV CD spectra (190-250 nm) of the protein were recorded in temperature intervals of 5°C from 20 to 90°C. The MRE (mean residue ellipticity) in deg·cm2·dmol–1 was calculated using the equation: MRE = θobs(mdeg)/10·n·l·Cp, where θobs is the CD in millidegree, n is the number of amino acid residues (212), l is the pathlength of the cell in cm (0.1), and Cp is the molar concentration of the protein.
The reversibility of temperature-induced changes was monitored by cooling the preheated samples from 90 to 20°C at a similar rate, i.e. 1°C/min. The percent α-helical and β-sheet contents of the protein under desired conditions were calculated using the online K2D3 software [23] with the MRE values from 190-240 nm as extracted from the CD spectra.
Turbidity measurements. The temperature-dependent changes (20-90°C) in turbidity of protein samples under desired conditions were observed by recording absorbance at 350 nm on a Perkin-Elmer Lambda 25 double beam UV-Vis spectrophotometer. The time-dependent changes in turbidity of the protein at different temperatures were also monitored in a similar manner.
The Rayleigh light scattering (RLS) experiment was performed using a Hitachi spectrofluorimeter (F-4500) in a 1 cm pathlength cuvette. Protein samples under the desired conditions were excited at 350 nm, and emission spectra were recorded from 300 to 400 nm. Both excitation and emission slits were fixed at 5 nm.
ANS fluorescence assay. A stock solution of ANS was prepared in distilled water, and its concentration was determined using molar extinction coefficient εM = 5000 M–1·cm–1 at 350 nm. For the ANS binding experiment, protein samples under different conditions were incubated with 50-fold molar excess of ANS for 30 min in the dark. The excitation wavelength for ANS fluorescence was set at 380 nm, and emission spectra were recorded from 400 to 600 nm. Both the excitation and emission slits were set at 5 nm.
Proteolytic activity assay. The proteolytic activity of papain at different temperatures was assayed using 1% casein solution as the substrate. For measuring the activity, the modified enzyme was incubated in the activation buffer (20 mM sodium phosphate, pH 7.0) containing 5 mM cysteine and 1 mM EDTA for 2 min. The reaction was initiated by adding 0.1 ml of papain (5 μM), which was preincubated at the desired temperature for 5 min to 0.9 ml of casein solution. The reaction was stopped after exactly 10 min by adding 1 ml of 10% trichloroacetic acid (TCA). The samples were allowed to stand for 30 min, followed by centrifugation at 3000g for 20 min. The absorbance of the filtered supernatant was recorded at 280 nm. Background hydrolyses of the substrate as well as enzyme were measured and used for correcting the absorbance values. Enzyme activity was reported as change in absorbance at 280 nm/min. The experiments were performed in triplicates.
Transmission electron microscopy. The morphology of aggregates under different conditions was examined using a JEOL 2100F transmission electron microscope (TEM) operating at accelerating voltage of 200 kV. The protein sample (6 µl), incubated under different conditions, was applied on 200 mesh copper grid covered by carbon film. Excess of fluid was removed after 2 min, the unbound protein was washed with Milli-Q water, and the samples were negatively stained with 2% (w/v) uranyl acetate. The grids then were air-dried and viewed under the electron microscope.
RESULTS
Monitoring the thermal unfolding of papain by far-UV CD. The thermal denaturation of papain in the temperature range 20-90°C was observed by the far-UV CD method, which reflects changes in the regular secondary structure of a protein [24]. Figure 1 shows a characteristic thermal unfolding profile of papain at three different concentrations, i.e. 3, 5, and 10 µM, displaying an almost similar trend. The negative ellipticity (θ) value at 222 nm remained almost unchanged between 20 to 50°C. Heating the samples above 50°C caused a gradual decrease in negative ellipticity, which indicates unfolding of the protein. The negative ellipticity was found to decrease significantly in between 70-80°C, which suggests a significant loss of regular secondary structure. No further changes in ellipticity were observed above 80°C.
Fig. 1. Temperature-dependent changes in ellipticity (θ) at 222 nm of papain at different concentrations. For sake of clarity, the reversibility profile is shown for only the 10 μM protein concentration.
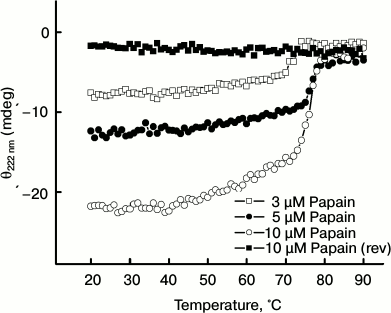
To check if temperature-induced changes in protein conformation are reversible or not, the samples were cooled from 90 to 20°C at a similar rate. However, no detectable increase in (negative) ellipticity was observed on cooling the preheated enzyme (10 µM), thus suggesting that the thermal unfolding of papain is an irreversible process. Similar results were observed at the other protein concentrations (data not shown for sake of clarity). From these results, it is clear that papain undergo an irreversible unfolding, which is in agreement with previous observations [20].
Thermal aggregation of papain as monitored by turbidity measurement. To assess the cause of irreversible unfolding of papain, we observed temperature-dependent changes in turbidity of the protein at different concentrations as shown in Fig. 2. The turbidity of protein samples remained insignificant in the temperature range 20-50°C. Upon raising the temperature above 50°C, a gradual increase in turbidity was observed. The turbidity was markedly enhanced at about 70-80°C. This indicates that unfolding of the protein (as seen in Fig. 1) is accompanied by simultaneous aggregation.
Fig. 2. Temperature-dependent changes in turbidity at 350 nm at different concentrations of papain.
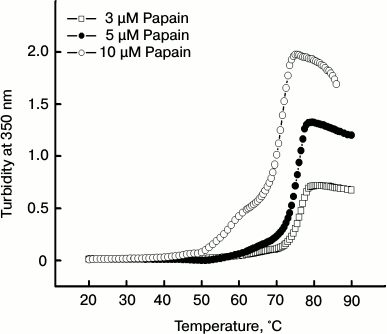
It is notable that turbidity of the samples increases with increasing concentration of the protein. Figure 3 shows a picture of papain samples at different concentrations that were subjected to 70°C for 1 h followed by overnight incubation. Increasing amounts of aggregate can be seen in samples containing 3-40 μM papain.
Fig. 3. Eppendorf tubes containing increasing concentrations of papain (pH 7.0) incubated at 70°C for 1 h and kept overnight.

Effect of SDS on thermal aggregation of papain. As mentioned earlier, post-micellar concentration of the anionic surfactant SDS is reported to exert inhibitory action on amyloid formation of some pathogenic proteins. To observe how SDS affects the thermal aggregation of papain, we incubated protein samples with 6 mM SDS. (The CMC of SDS in sodium phosphate buffer at neutral pH (25°C) is reported to be 4.7 mM as determined by conductivity experiments [25].)
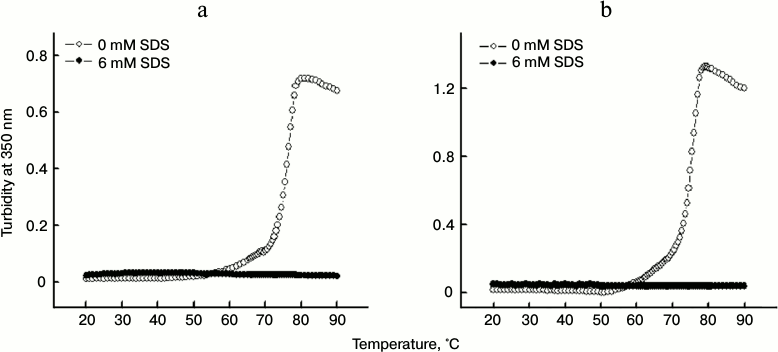
The change in turbidity of the protein in the absence and presence of SDS were examined for 3 and 5 µM protein samples (Figs. 4a and 4b, respectively) in the temperature range 20-90°C. Whereas protein samples incubated without SDS showed significant increase in turbidity when subjected to thermal treatment, the samples treated with 6 mM SDS did not exhibited any change in the turbidity over the entire temperature range studied. This suggested that SDS inhibits the thermal aggregation of papain, thereby not allowing the turbidity to rise. Furthermore, it was found that increasing the protein concentration from 3 to 5 μM did not affect the inhibitory action of SDS at the concentration taken in the study (6 mM). Similar results were obtained at higher a protein concentration (data not shown).
Fig. 4. Temperature-dependent changes in turbidity of 3 μM (a) and 5 μM (b) papain in the absence and presence of 6 mM SDS.
Effect of SDS on time-dependent changes in aggregation of papain. We also observed time-dependent changes in turbidity of papain at different temperatures in the absence and presence of SDS. The experiments were conducted at 3 μM (Fig. 5a) and 5 μM (Fig. 5b) protein concentration. The turbidity of samples increased with increase in time such that at 65 and 70°C no slope off was observed even at 30 min. However at 75°C very significant increase in turbidity was noticed within the first 5 min of incubation. After this the turbidity ceased to enhance further. A slight decrease in turbidity observed after 10 min at 75°C (at both the concentrations taken) may be attributed to the formation of large amount of aggregates that might have settled in the cuvette.
Fig. 5. Time-dependent changes in thermal aggregation of 3 μM (a) and 5 μM (b) papain at different temperatures in the absence and presence of 6 mM SDS.
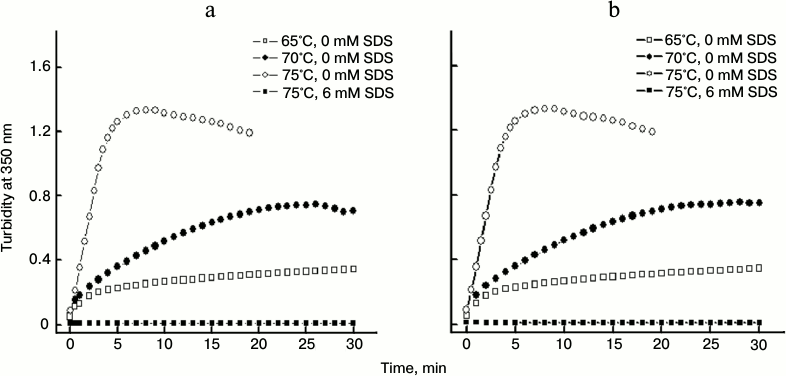
A similar interpretation may be true for the sample containing 5 μM protein at 70°C after about 25 min. In the presence of 6 mM SDS for 15 min prior to incubation at 75°C, no change in turbidity could be observed throughout the time period studied. These results clearly demonstrate the strong potential of post-micellar SDS concentration to inhibit thermal aggregation of papain.
Effect of SDS on thermal unfolding of papain. The remarkable potential of SDS in inhibiting the aggregation of the protein prompted us to examine how this surfactant affects the thermal unfolding of papain. For this, we followed the temperature-dependent changes in ellipticity of papain in the absence and presence of 6 mM SDS as shown in Fig. 6. In the absence of SDS, papain showed a characteristic thermal unfolding profile where the negative MRE at 222 nm decreased significantly from –11,739 deg·cm2·dmol–1 at 20°C to –3347 deg·cm2·dmol–1 at 90°C. On the other hand, in the presence of 6 mM SDS, the (negative) MRE value underwent only a moderate change from –10,654 deg·cm2·dmol–1 at 20°C to –7752 deg·cm2·dmol–1 at 90°C. This indicates that SDS may tend to suppress the unfolding of the protein, whereby its hydrophobic patches are not exposed enough to commence the aggregation process.
Fig. 6. Temperature-dependent changes in MRE222 nm of papain (5 μM) in the absence and presence of 6 mM SDS.
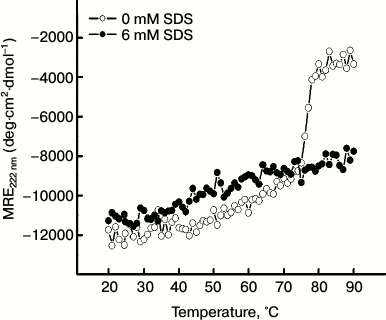
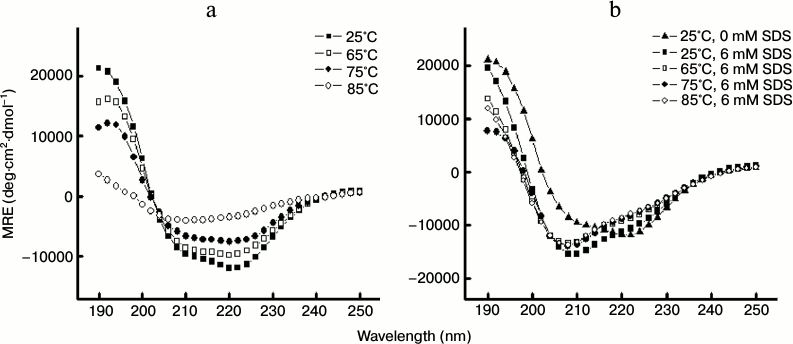
To assess the temperature-induced changes in protein conformation in the absence and presence of SDS, we collected far-UV CD spectra of the protein in different temperature intervals during the thermal denaturation experiments (as described in “Materials and Methods”, section “Circular dichroism”).
As evident from Fig. 7a (in the absence of the detergent), the far-UV CD spectra of papain at 25°C exhibited negative peaks at 208 and 222 nm, which are characteristics of α-helical content. As the temperature increased to 65 and to 75°C, we obtained spectra with considerably reduced negative ellipticity values suggesting loss of regular secondary structure. When the temperature was further raised to 85°C, the negative peaks at 208 and 222 nm almost vanished. On the other hand, CD spectra in the presence 6 mM SDS showed signs of induction, rather than loss of regular secondary structure, which persisted even at higher temperatures. This was inferred from a distinguishable increase in negative MRE at 208 nm at 25°C in the presence of 6 mM SDS as shown in Fig. 7b. The change in the spectra may be attributed to increase in helical content of the protein. Raising the temperature up to 85°C resulted in only marginal changes in the spectra without any significant loss of secondary structure.
Fig. 7. Far-UV CD spectra of papain (5 μM) at different temperatures in the absence (a) and presence (b) of 6 mM SDS. The samples were incubated for 1 h prior to monitoring the CD spectra.
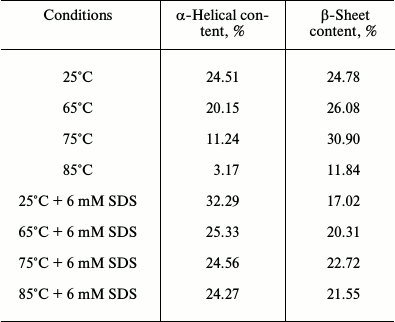
The table reveals changes in the percent α-helical and β-sheet contents of papain under different conditions. At 25°C, the protein was found to contain 24.51% α-helical and 24.78% β-sheet structures, which is similar to that obtained from the crystal structure of the enzyme (25% helical and 21% β-sheet structure) [26]. When the temperature was raised to 65°C, the α-helical content was reduced to 20.15% while β-sheet content was slightly enhanced to 26.08%. Increasing the temperature to 75°C led to a considerable lowering of α-helical content to 11.24%. On the other hand, β-sheet content was found to remain further enhanced (30.9%). This may be due to the fact that the two domains of papain, i.e. the α-domain and the β-domain, are known to fold independently, as mentioned earlier [20]. It is possible that unfolding the α-domain may have resulted in a conformation with β-sheet-rich structure that might have facilitated the aggregation reaction. At 85°C we observed that both α-helical and β-sheet content were drastically reduced, suggesting a significant loss of regular secondary structural elements.
In contrast, when the protein samples were incubated with 6 mM SDS prior to thermal unfolding, a marked increase in α-helical content (32.29%) was noted at 25°C, whereas the β-sheet content was reduced to 17.02%. At higher temperatures, i.e. 65-85°C, the α-helical content was moderately reduced but not as significantly lost as in the absence of SDS. Besides, β-sheet content also did not alter as radically as during the aggregation of the protein. These data clearly show that the post-micellar SDS concentration enhances the helical propensity of papain, which may be responsible for the observed inhibition of thermal aggregation of the protein.
Changes in % α-helical and β-sheet contents of papain
(pH 7.0) under different conditions
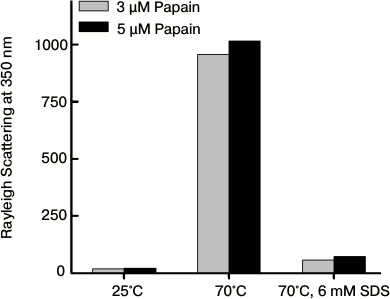
Monitoring the thermal aggregation of papain by the Rayleigh scattering method. So far we have determined the thermal aggregation of papain only by turbidity measurements. Another way to analyze the presence of aggregates is to monitor the light scattering intensity [15]. Figure 8 shows light scattering intensity at 350 nm of papain at 3 and 5 µM after being subjected to 25 and 70°C in the absence of SDS and also at 70°C in the presence of 6 mM SDS. At 25°C, papain showed very insignificant light scattering intensity indicating the absence of aggregates in the samples. Incubating the samples at 70°C for 1 h markedly increased the light scattering intensity, which suggests formation of a large amount of aggregates. However, protein samples at 70°C that were pretreated with 6 mM SDS exhibited little scattering intensity, which was comparable to that observed at 25°C. It is also notable that the light scattering intensity is greater for the sample containing 5 µM papain compare to 3 µM papain. The data further confirm that papain undergoes thermal aggregation, the extent of which increases with increase in protein concentration. Furthermore, SDS at the post-micellar concentration efficiently inhibited its thermal aggregation.
Fig. 8. Light scattering intensity at 350 nm in papain samples at different temperatures in the absence and presence of 6 mM SDS.
Thermal aggregation of papain as studied by ANS fluorescence. We also employed extrinsic fluorescence assay to further determine the aggregation of protein by using ANS as an extrinsic fluorophore [27]. The preferential binding of ANS to hydrophobic patches gives rise to enhanced fluorescence emission accompanied by a blue shift of the spectral maximum, and therefore the surface hydrophobicity of protein samples (which is enhanced during aggregation) can be monitored by ANS fluorescence emission [28, 29].
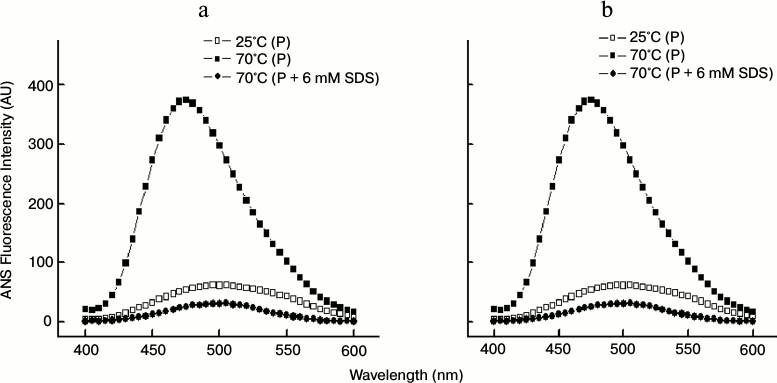
As shown in Figs. 9a and 9b, at 25°C the ANS fluorescence spectra of 3 and 5 µM papain, respectively, exhibited very low fluorescence intensity with an emission maxima (λmax) at 505 nm indicating that the hydrophobic sites are hidden inside the well-folded native papain. Incubation of the protein at 70°C clearly augmented the ANS fluorescence intensity at 480 nm along with a blue-shifted λmax, thus further confirming the formation of aggregates. In contrast, the protein samples at 70°C that were preincubated with 6 mM SDS gave very low fluorescence intensity (spectra obtained after subtracting from appropriate controls), thus indicating suppression of aggregation.
Fig. 9. ANS fluorescence spectra of 3 μM (a) and 5 μM (b) papain at different temperatures in the absence and presence of 6 mM SDS.
Morphology of thermally induced aggregates of papain as examined by TEM. The morphology of aggregates of papain under different conditions was examined using transmission electron microscopy as shown in Fig. 10. It can be observed that aggregates with amorphous, not fibrillar, appearance were found in 3 and 5 µM papain-containing samples (Figs. 10a and 10b, respectively) that were incubated at 70°C for 1 h prior to imaging. Also, the aggregates formed were denser at the higher concentration. On the other hand, protein samples treated with 6 mM SDS (Figs. 10c and 10d) revealed almost negligible aggregate formation. It is also evident from the TEM images that thermally induced aggregates of papain lacked the characteristics of typical amyloid structures.
Fig. 10. Transmission electron microscopic images of: 3 μM papain in the absence (a) and presence (c) of 6 mM SDS; 5 μM papain in the absence (b) and presence (d) of 6 mM SDS. The scale bar corresponds to 100 nm.

Effect of SDS concentration on the activity of papain. As mentioned earlier, the fact that a post-micellar SDS concentration effectively inhibits aggregation of the protein can be exploited as a useful approach only if the protein retains its activity in the presence of SDS. Therefore, the activity of papain was examined with respect to temperature in the absence and presence of 6 mM SDS. The activity of papain was determined using casein as the substrate. The synthetic substrates that are more commonly employed for activity measurements could not be used because they were either precipitated by SDS (Nα-benzoyl-L-arginine 4-nitroanilide hydrochloride, BAPNA) or they were themselves undergoing autolysis at pH 7.0 (Z-L-Lys-ONp hydrochloride). It should also be noted that adequate controls of substrate casein were taken to rule out its chaperone activity and changes induced by casein on protein conformation. Figure 11a shows the activity of papain at different temperatures in the absence and presence of 6 mM SDS. The percent (%) activity was calculated taking the activity of the enzyme at 65°C (where maximum activity was recorded) as 100%. It is evident from this figure that papain remains considerably active over a broad range of temperature 35-75°C, in accordance with previous reports. In contrast, the enzyme showed no appreciable activity in the presence of SDS. Only 8 to 12% of the activity was retained above 50°C in the presence of 6 mM SDS. These findings clearly disfavor the use of SDS as a suitable aggregation inhibitor for papain.
Fig. 11. a) Temperature-dependent changes in activity of papain in the absence and presence of 6 mM SDS. b) Effect of 5 mM cysteine and 1 mM EDTA on thermal aggregation of papain at 70°C.
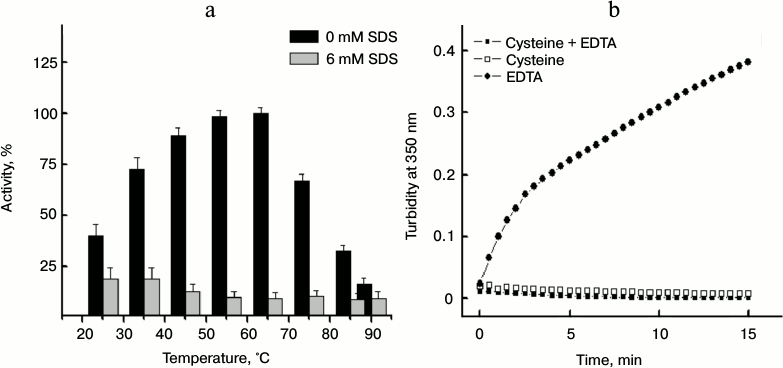
The activity measurement data shown above raise a question: how is the protein showing activity under conditions where it is found to aggregate? The key to this question is that prior to the activity measurements the enzyme was incubated in the activation buffer containing cysteine and EDTA as mentioned in “Materials and Methods” section. In Fig. 11b, we showed the time-dependent changes in turbidity at 350 nm of papain at 70°C in buffer containing either 5 mM cysteine or 1 mM EDTA or both. From this figure it is clear that cysteine, but not EDTA, protects the enzyme against thermal aggregation, thereby facilitating activity measurements. The results obtained from activity measurements also reveal that despite being incubated in activation buffer (containing cysteine), papain is not able to retain its activity in the presence of SDS.
DISCUSSION
Protein aggregation remains one of the major challenges in the development and commercialization of biotechnology products. Among all the environmental factors that determine aggregation of proteins, temperature is arguably the most critical factor for consideration when proteins are handled during the entire development and commercialization processes [3]. It is reported that the thermodynamic stability of protein resides over only a narrow temperature range [30]. Therefore, decreasing or increasing the temperature may destabilize a protein and can promote aggregation. Surfactants are often added into protein solutions to inhibit protein aggregation in different processes, such as shaking/shearing as well as thermal treatment. Their stabilization effect has been mostly attributed to their weak binding to proteins, thus blocking or partially blocking the aggregation-prone hydrophobic sites on the protein surface [31]. Additionally, protein–surfactant interaction holds considerable importance due to its application in pharmaceutical, chemical, and cosmetic industries [1, 32]. Ionic surfactant, particularly SDS, has been demonstrated to effectively inhibit the self-assembly of protein above its CMC.
In the present study, we demonstrate that papain, a well-utilized enzyme of great industrial and medicinal importance, undergoes thermal aggregation. Initially, the thermal unfolding of papain was monitored at different concentrations by far-UV CD. We observed that the protein significantly unfolds between 50-80°C. In addition, the thermal melting of this protein was found to be an irreversible process. To explore the cause of irreversible unfolding, the turbidity of the protein was monitored with respect to temperature, and it was observed that the protein unfolding above 50°C is accompanied by a simultaneous aggregation process. This thermal aggregation behavior of papain was observed over a concentration range of 3-40 µM. The reason for analyzing the effect of protein concentration on thermal aggregation is the existence of different aggregation pathways due to which increase in protein concentration may result in different consequences, such as decreased aggregation due to crowding effect or increased aggregation due to increased chance of association or precipitation due to the solubility limit. The ultimate effect is protein-dependent and the net result of these factors [3].
Aggregation was also confirmed by increase in light scattering intensity and enhancement of ANS fluorescence intensity. The morphology of the aggregates was examined by transmission electron microscopy, where aggregates with disordered or amorphous appearance were observed. Furthermore, we monitored the influence of post-micellar SDS concentration on aggregation of papain. It was found that SDS potentially inhibits the self-assembly of papain as demonstrated by insignificant turbidity, reduced light scattering intensity, and significantly lowered ANS fluorescence intensity. Besides, TEM images of the protein incubated with post-micellar SDS concentration did not reveal any aggregated morphology.
To determine the cause of aggregation inhibition by SDS, the far-UV CD spectra of the protein were collected at different temperatures in the absence and presence of SDS. The CD spectra of the protein in the absence of SDS showed a normal unfolding behavior where negative ellipticity values continuously decreased with increase in temperature. However, in the presence of SDS, induction of helical structure was observed. The calculated percent α-helical content was found to decrease from 24% at 25°C to nearly 3% at 85°C in the absence of SDS. However, incubation with 6 mM SDS at 25°C led to an increase in α-helical content to 32%. Heating to 85°C led to only a moderate decrease (to 24%) in helicity. These observations led us to conclude that increased helical content as induced by SDS might have stabilized papain against thermal denaturation. Similar conclusions have been drawn in previous studies on lysozyme, insulin, α-synuclein, etc., where aggregation inhibition at post-micellar SDS concentration has also been attributed to increase in helical structure contents of the respective model proteins [14, 16, 17]. Additionally, in a recent study on thermal unfolding of bovine serum albumin (BSA), it was shown that the helicity of BSA decreases sharply between 50-100°C, but in the presence of SDS the helicity was remarkably maintained up to 85°C. The protective effect of SDS on secondary structure of BSA and other serum albumins (which mainly contain helical structure) is observed not only during thermal denaturation, but also during the chemical denaturation process [33].
The strong aggregation inhibition potential of SDS prompted us to check the activity of papain in the presence of this inhibitor and to examine whether such an approach can be successfully applied to proteins that are unrelated to pathology but hold immense commercial importance. However, the results obtained from activity measurements clearly showed papain to be functionally inactive when incubated with post-micellar SDS concentration. A nearly 10-fold decrease in enzyme activity was observed in the presence of 6 mM SDS above 50°C. From these findings, it appears that SDS might have protected the secondary structure of the protein but strongly perturbed its tertiary conformation, which is crucial for the enzyme activity.
Keeping in consideration all the above-described facts, it can be said that the conformational alterations brought about by SDS were indeed making papain structurally stable but rendering it functionally inactive. The overall results of this study are summarized in a schematic form as shown in Fig. 12.
Fig. 12. Schematic representation of temperature-induced changes in papain conformation in the absence and presence of SDS (>CMC).
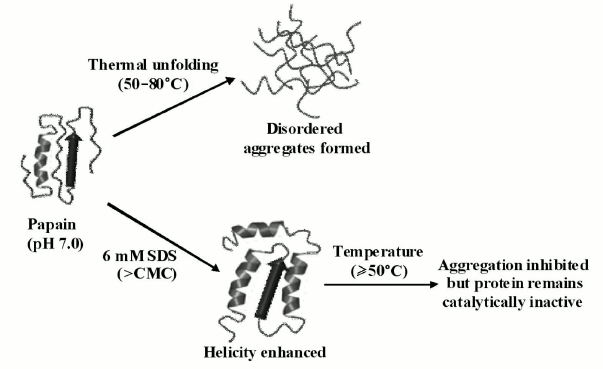
In conclusion, we observed that papain exhibits remarkable tendency to aggregate above 50°C. The aggregation propensity increases with increases in protein concentration. The aggregates formed were disordered in appearance and lacked characteristics of amyloid fibrils. Furthermore, we checked the effect of post-micellar SDS concentration on the self-assembly process and found that SDS significantly inhibits the aggregation of papain by inducing the formation of helical structure whereby the protein resisted unfolding at higher temperatures and therefore does not aggregate. Unfortunately, despite being an effective aggregation inhibitor, SDS was found to be unable to restore the activity of the enzyme. With these facts in consideration, we conclude that promoting the helical propensity of protein to attain aggregation inhibition may not be a successful strategy to overcome the aggregation-related problems of enzymes such as papain.
Facilities provided by the Interdisciplinary Biotechnology Unit, AMU, Aligarh are gratefully acknowledged. We are thankful to the Council of Scientific and Industrial Research (CSIR) for providing Senior Research Fellowship to AQ.
REFERENCES
1.Wang, W. (2005) Protein aggregation and its
inhibition by biopharmaceutics, Int. J. Pharm., 289,
1-30.
2.Kudou, M., Shiraki, K., Fujiwara, S., Imanaka,
T., and Takagi, M. (2003) Prevention of thermal inactivation and
aggregation of lysozyme by polyamines, Eur. J. Biochem.,
270, 4547-4554.
3.Wang, W., Nema, S., and Teagarden, D. (2010)
Protein aggregation-pathways and influencing factors, Int. J.
Pharm., 390, 89-99.
4.Rondeau, P., Navarra, G., Cacciabaudo, F.,
Leone, M., Bourdon, E., and Militello, V. (2010) Thermal
aggregation of glycated bovine serum albumin, Biochim. Biophys.
Acta, 1804, 789-798.
5.Belton, D. J., and Miller, A. F. (2013) Thermal
aggregation of recombinant protective antigen: aggregate morphology and
growth rate, J. Biophys., 75, 1091.
6.Mession, J. L., Sok, N., Assifaoui, A., and Saurel,
R. (2013) Thermal denaturation of pea globulins (Pisum sativum
L.) – molecular interactions leading to heat-induced protein
aggregation, J. Agric. Food Chem., 61, 1196-1204.
7.Fallah-Bagheri, A., Moosavi-Movahedi, A. A.,
Taghizadeh, M., Khodarahmi, R., Ma’mani, L., Bijari, N.,
Bohlooli, M., Shafiee, A., Sheibani, N., and Saboury, A. A. (2013)
Modified β-casein restores thermal reversibility of human carbonic
anhydrase II: salt bridge connection, Biotechnol. Appl.
Biochem., 60, 298-304.
8.Yan, Y. B., Wang, Q., He, H. W., and
Zhou, H. M. (2004) Protein thermal aggregation involves distinct
regions: sequential events in the heat-induced unfolding and
aggregation of hemoglobin, Biophys. J., 86,
1682-1690.
9.Militello, V., Casarino,
C., Emanuele, A., Giostra, A., Pullara, F., and
Leone, M. (2004) Aggregation kinetics of bovine serum albumin
studied by FTIR spectroscopy and light scattering, Biophys.
Chem., 107, 175-187.
10.Vetri, V., Librizzi, F., Leone, M., and
Militello, V. (2007) Thermal aggregation of bovine serum albumin
at different pH: comparison with human serum albumin, Eur. Biophys.
J., 36, 717-725.
11.Das, U., Hariprasad, G., Ethayathulla,
A. S., Manral, P., Das, T. K., Pasha, S., Mann,
A., Ganguli, M., Verma, A. K., Bhat,
R., Chandrayan, S. K., Ahmed, S., Sharma, S., Kaur,
P., Singh, T. P., and Srinivasan, A. (2007) Inhibition of
protein aggregation: supramolecular assemblies of arginine hold the
key, PLoS One, 2, e1176.
12.Hamada, H., Arakawa, T., and Shiraki,
K. (2009) Effect of additives on protein aggregation, Curr. Pharm.
Biotechnol., 10, 400-407.
13.Pertinhez, T.
A., Bouchard, M., Smith, R. A., Dobson, C.
M., and Smith, L. J. (2002) Stimulation and inhibition of fibril
formation by a peptide in the presence of different concentrations of
SDS, FEBS Lett., 529, 193-197.
14.Hung, Y. T., Lin, M. S., Chen, W.
Y., and Wang, S. S. (2010) Investigating the effects of sodium
dodecyl sulfate on the aggregative behavior of hen egg-white lysozyme
at acidic pH, Colloids Surf. B. Biointerfaces, 81,
141-151.
15.Khan, J. M., Qadeer, A., Chaturvedi, S. K.,
Ahmad, E., Rehman, S. A., Gourinath, S., and Khan, R. H. (2012) SDS can
be utilized as an amyloid inducer: a case study on diverse proteins,
PLoS One, 7, e29694.
16.Giehm, L., Oliveira, C.
L., Christiansen, G., Pedersen, J. S., and Otzen, D. E.
(2010) SDS-induced fibrillation of alpha-synuclein: an alternative
fibrillation pathway, J. Mol. Biol., 401, 115-133.
17.Badraghi, J., Moosavi-Movahedi, A.
A., Saboury, A. A., Yousefi, R., Sharifzadeh,
A., Hong, J., Haertle, T., Niasari-Naslaji, A., and
Sheibani, N. (2009) Dual behavior of sodium dodecyl sulfate as enhancer
or suppressor of insulin aggregation and chaperone-like activity of
camel alpha-S(1)-casein, Int. J. Biol. Macromol., 45,
511-517.
18.Edwin, F., and Jagannadham, M. V. (1998)
Sequencing unfolding of papain in molten globule state, Biochem.
Biophys. Res. Commun., 252, 654-660.
19.Grzonka, Z., Kasprzykpwski, F., and Wiczk, W.
(2007) in Industrial Enzymes: Structure, Function and
Applications (Polaina, J., and MacCabe, A. P., eds.) Springer, pp.
181-195.
20.Solis-Mendiola, S., Rojo-Dominguez,
A., and Hernandez-Arana, A. (1993) Cooperativity in the unfolding
transitions of cysteine proteinases. Calorimetric study of the heat
denaturation of chymopapain and papain, Biochim. Biophys.
Acta, 1203, 121-125.
21.Fatima, S., and Khan, R. H. (2007) Effect of
polyethylene glycols on the function and structure of thiol proteases,
J. Biochem., 142, 65-72.
22.Mitchel, R. E., Chaiken, I. M., and
Smith, E. L. (1970) The complete amino acid sequence of papain.
Additions and corrections, J. Biol. Chem., 245,
3485-3492.
23.Louis-Jeune, C., Andrade-Navarro, M.
A., and Perez-Iratxeta, C. (2011) Prediction of protein secondary
structure from circular dichroism using theoretically derived spectra,
Proteins, 80, 374-381.
24.Kelly, S. M., Jess, T. J., and Price, N. C.
(2005) How to study proteins by circular dichroism, Biochim.
Biophys. Acta, 1751, 119-139.
25.Rangel-Yagui, C. O., Hsu, H. W. L., Pessoa, A.,
Jr., and Tavares, L. C. (2005) Micellar solubilization of ibuprofen
– influence of surfactant head groups on the extent of
solubilization, Brazil. J. Pharm. Sci., 41, 237-246.
26.Kamphuis, I. G., Kalk, K. H., Swarte,
M. B., and Drenth, J. (1984) Structure of papain refined at 1.65 A
resolution, J. Mol. Biol., 179, 233-256.
27.Hawe, A., Sutter, M., and Jiskoot,
W. (2008) Extrinsic fluorescent dyes as tools for protein
characterization, Pharm. Res., 25, 1487-1499.
28.McDuff, F. O., Doucet, A., and
Beauregard, M. (2004) Low concentration of guanidine hydrochloride
induces the formation of an aggregation-prone state in alpha-urease,
Biochem. Cell Biol., 82, 305-313.
29.Morshedi, D., Rezaei-Ghaleh,
N., Ebrahim-Habibi, A., Ahmadian, S., and
Nemat-Gorgani, M. (2007) Inhibition of amyloid fibrillation of
lysozyme by indole derivatives – possible mechanism of action,
FEBS J., 274, 6415-6425.
30.Talla-Singh, D., and Stites, W. E. (2008)
Refinement of noncalorimetric determination of the change in heat
capacity, ΔCp, of protein unfolding and validation
across a wide temperature range, Proteins, 71,
1607-1616.
31.Mahler, H. C., Friess, W., Grauschopf,
U., and Kiese, S. (2009) Protein aggregation: pathways, induction
factors and analysis, J. Pharm. Sci., 98,
2909-2934.
32.Li, Y., Cao, M., and Wang, Y.
(2006) Alzheimer amyloid beta(1-40) peptide: interactions with cationic
gemini and single-chain surfactants, J. Phys. Chem. B,
110, 18040-18045.
33.Moriyama, Y., Watanabe, E., Kobayashi,
K., Harano, H., Inui, E., and Takeda, K. (2008)
Secondary structural change of bovine serum albumin in thermal
denaturation up to 130°C and protective effect of sodium dodecyl
sulfate on the change, J. Phys. Chem. B, 112,
16585-16589.