Plasticity of Central and Peripheral Sources of Noradrenaline in Rats during Ontogenesis
N. S. Bondarenko1, L. K. Dilmukhametova1, A. Yu. Kurina1, A. R. Murtazina1*, A. Ya. Sapronova1, A. P. Sysoeva1, and M. V. Ugrumov1,2
1Koltzov Institute of Developmental Biology, Russian Academy of Sciences, 119334 Moscow, Russia; E-mail: aliya_mr89@mail.ru2National Research University Higher School of Economics, 101000 Moscow, Russia
* To whom correspondence should be addressed.
Received November 16, 2016; Revision received December 2, 2016
The morphogenesis of individual organs and the whole organism occurs under the control of intercellular chemical signals mainly during the perinatal period of ontogenesis in rodents. In this study, we tested our hypothesis that the biologically active concentration of noradrenaline (NA) in blood in perinatal ontogenesis of rats is maintained due to humoral interaction between its central and peripheral sources based on their plasticity. As one of the mechanisms of plasticity, we examined changes in the secretory activity (spontaneous and stimulated release of NA) of NA-producing organs under deficiency of its synthesis in the brain. The destruction of NA-ergic neurons was provoked by administration of a hybrid molecular complex – antibodies against dopamine-β-hydroxylase associated with the cytotoxin saporin – into the lateral cerebral ventricles of neonatal rats. We found that 72 h after the inhibition of NA synthesis in the brain, its spontaneous release from hypothalamus increased, which was most likely due to a compensatory increase of NA secretion from surviving neurons and can be considered as one of the mechanisms of neuroplasticity aimed at the maintenance of its physiological concentration in peripheral blood. Noradrenaline secretion from peripheral sources (adrenal glands and the organ of Zuckerkandl) also showed a compensatory increase in this model. Thus, during the critical period of morphogenesis, the brain is integrated into the system of NA-producing organs and participates in their reciprocal humoral regulation as manifested in compensatory enhancement of NA secretion in each of the studied sources of NA under specific inhibition of NA production in the brain.
Keywords: noradrenaline, brain, neuroplasticity, adrenal glands, organ of Zuckerkandl, neonatal, ratsDOI: 10.1134/S0006297917030166
Abbreviations: anti-DBH–saporin, anti-dopamine-β-hydroxylase–saporin complex; BBB, blood–brain barrier; DBH, dopamine-β-hydroxylase; HPLC-ED, high performance liquid chromatography with electrochemical detection; LC, locus coeruleus; NA, noradrenaline; OZ, organ of Zuckerkandl.
The morphogenesis of organs and the organism as a whole occurs under the
control of intercellular chemical signals in the critical period of
ontogenesis, which corresponds mainly to the perinatal period of
development in rodents. One of the morphogenetic factors is
noradrenaline (NA), which is involved in the regulation of development
of the brain and peripheral target organs such as heart, blood vessels,
lungs, and lymphoid organs [1-4]. Impairment of NA metabolism in the perinatal
period results in irreversible, often fatal, developmental disorders
[3-5]. NA is synthesized
already in the middle of the prenatal period by neurons of the central
and peripheral nervous systems and chromaffin cells of the adrenal
glands and paraganglia [6-8].
Previously, we formulated and proved a hypothesis according to which the brain, before the formation of the blood–brain barrier (BBB), i.e. before the 2nd week of life in rats [9], is a source of NA in the general circulatory system [10, 11]. After the formation of the BBB, its secretion from the brain into blood ceases. Chromaffine cells of the medullary substance of the adrenal glands secrete NA through the lifespan, and their functional activity increases in ontogeny [12]. The organ of Zuckerkandl (OZ) – the largest assembly of extra-adrenal chromaffin cells in the area of bifurcation of the abdominal aorta – reaches its maximum activity at the end of the prenatal period and undergoes involution by the end of the second week of life [13]. All these organs in the studied period of ontogeny can be a source of NA in the general circulatory system [10, 11, 14]. However, NA concentration in peripheral blood is maintained at a level comparable with its level in the portal circulatory system of adult animals that is sufficient for regulation of the functions of the target organs [10-15].
The results of our previous studies [10, 11] suggest that the physiologically active concentration of NA in blood plasma in the perinatal period of ontogeny is maintained due to humoral interaction between its central and peripheral sources based on their plasticity. Such interaction also seems to be possible because the expression of adrenoceptor genes has been revealed in nerve and chromaffin cells of rats already in the prenatal period of development [16]. The enhanced secretory activity of some NA sources against reduced NA synthesis by other sources seems to be one of the potential mechanisms of plasticity.
This work was aimed at assessing the secretory activity of peripheral NA-producing organs against the decrease in its production in rat brain in the early postnatal period. The research objectives were as follows: (i) morphological assessment of the NA-ergic system of the brain during its specific destruction; (ii) investigation of changes in spontaneous and stimulated NA secretion from brain (hypothalamus), adrenal glands, and OZ in case of impaired NA production in the brain.
MATERIALS AND METHODS
Animals. Male Wistar rats were used in the work on day 2, 4, and 5 of life. The birthday of rats was considered as the first day of life. The rats were kept under standard vivarium conditions with free access to food and water.
Chronic inhibition of noradrenaline synthesis in rat brain. On day 2 of life, the rats were anaesthetized with isoflurane, the skin on the head was dissected, and a hole in the skull (2 × 2 mm) was cut out laterally from the bregma. Then the animals were placed into a stereotaxic device adapted for young animals, and a glass microcannula (50 µm in diameter) connected through a Teflon tube to a Hamilton microsyringe was introduced into the lateral ventricle of brain by the following coordinates: 1.2 mm laterally from the bregma, 2.5 mm deep into the brain. For destruction of NA-ergic neurons, young rats were administered with anti-dopamine-β-hydroxylase–saporin (anti-DBH–saporin) – a hybrid molecular complex consisting of antibodies against dopamine-β-hydroxylase (DBH) associated with the cytotoxin saporin [17, 18]. In the experiment (n = 10), 0.5 µg of anti-DBH–saporin in 2 µl of 0.9% NaCl was introduced stereotaxically into the lateral ventricle of the brain. In the control, 2 µl of 0.9% NaCl was introduced.
Immunohistochemical detection of dopamine-β-hydroxylase in rat brain. Forty-eight hours after the introduction of anti-DBH–saporin (n = 4) or 0.9% NaCl (n = 4), 0.02 M PBS (pH 7.2-7.4) was perfused through the heart of anaesthetized animals for 10 min at 37°C; then 4% paraformaldehyde in 0.2 M phosphate buffer was perfused for 10 min at 4°C. The animals were decapitated. The brain was extracted, additionally fixed by immersion in 4% paraformaldehyde for 12 h at 4°C, washed three times in PBS for 10 min, incubated in 20% sucrose solution in PBS for 24 h at 4°C, frozen in hexane cooled to –40°C, and stored at –70°C.
For DBH immunolabeling with peroxidase in a Leica CM1950 cryostat (Leica, Germany), serial frontal sections of the brainstem were made at the level of the locus coeruleus (LC), 14 µm thick (from the bregma, 5.20; before the bregma, 6.00) [19, 20]. Each second LC section from one control and one experimental animal was mounted on one slide. Then the sections on the slides were successively incubated in 0.02 M PBS solutions (pH 7.2-7.4): (i) with 3% BSA (Sigma, USA) and 0.1% Triton X-100 (Sigma) for 30 min at 20°C; (ii) with murine monoclonal antibodies to dopamine-β-hydroxylase (diluted 1 : 300) (Millipore, USA) in 3% BSA and 0.1% Triton X-100 for 36 h at 4°C; (iii) with biotinylated goat antibodies against mouse immunoglobulins (diluted 1 : 200) (Vector Laboratories, USA) for 2 h at 20°C; and (iv) with avidin–biotin complex associated with horseradish peroxidase (Vector Laboratories) for 1 h at 20°C. The sections were washed in PBS three times for 15 min each after all incubations (except for the first one), and three times for 30 min each after the last incubation. The avidin–biotin complex peroxidase was detected by incubation with 0.05% 3,3-diaminobenzidine tetrahydrochloride (Sigma) and 0.02% H2O2 in PBS at 20°C under visual control of all slides simultaneously. Then the sections were dehydrated in alcohol series of increasing concentrations, clarified in toluene, and embedded in Permount DePeX (Sigma).
The LC sections after DBH immunolabeling were examined in an Olympus BX51 light microscope (Olympus, Japan) with lens magnification of ×20/0.5 and photographed with an Olympus DP70 digital camera (Olympus). The number of DBH-immunoreactive neurons in the left part of the LC was counted on the section images using AnalySIS 5.0 software (Olympus).
Flow incubation of hypothalamus, adrenal glands, and organ of Zuckerkandl. The brain, adrenal glands, and OZ were isolated 72 h after the intraventricular introduction of anti-DBH–saporin (n = 6) or 0.9% NaCl (n = 6) on day 2 of life of the rats. Then the whole hypothalamus was separated from the brain, and 300-µm frontal sections were made with a Vibratome Series 1000 (Intracel Ltd., GB) in Krebs–Ringer solution containing NaCl, 120 mM; KCl, 4.8 mM; CaCl2, 2 mM; MgSO4, 1.3 mM; NaHCO3, 25 mM; D-glucose, 10 mM; HEPES, 20 mM; ascorbic acid, 0.1 mM (pH 7.2-7.4, 4°C). The adrenal glands were cut into four parts; the OZ was cut into halves. The sections of the hypothalamus and peripheral organs were placed into 400-µl thermostatic (37°C) flow chambers and incubated in the Krebs–Ringer solution for 1 h, the solution flowing through the chamber at a rate of 90 µl/min. Then three successive 10-min fractions of the incubation medium were collected (spontaneous secretion). Krebs–Ringer solution with a high content of K+ (56 mM) was fed into the chambers for 10 min, and then one 10-min fraction was collected (K+-stimulated secretion). Solutions of 1 M HClO4 (150 µl) and 3,4-dihydroxybenzylamine (1-12.5 pmol depending on the type of tissue and the age of animal) were added to the collected fractions of the medium as an internal standard required for further determination of NA by high performance lipid chromatography with electrochemical detection (HPLC-ED).
The average weight of tissue pieces used in the flow incubation of the hypothalamus, adrenal glands, and OZ was the same in the control and in the experiment.
High performance liquid chromatography with electrochemical detection (HPLC-ED). The tissues were homogenized in 10 volumes of 0.1 M HClO4 containing 25-250 pmol/ml of 3,4-dihydroxybenzylamine with a Labsonic M homogenizer (Sartorius AG, Germany) and centrifuged at 2000g for 20 min. For determination of NA in the incubation medium and cerebral tissue, the supernatant was supplemented with buffer (Tris-EDTA, pH 8.6) and extracted on aluminum oxide. NA was measured by reversed-phase HPLC-ED (Shimadzu LC 20ADSP with an Antec Decade II detector) at potential 850 mV [21] in a C18 Reprosil-Pur ODS-3 column, 3 µm, 100 × 4 mm (Dr. Maisch, Germany). The injector loop volume was 25 µl. The F-test for sample homogeneity assessment and the Student’s test for determination of reliability of differences were used for statistical data processing. Both tests are included in Sigma Plot Version 9.1 and GraphPad Prism Version 5.0 software packages for Windows (GraphPad Software, USA).
RESULTS
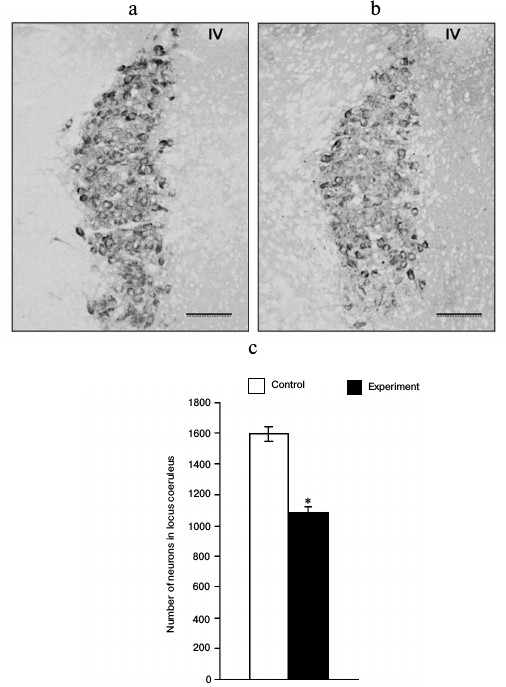
Immunohistochemical characteristics of noradrenergic neurons of brain after introduction of anti-DBH–saporin. The number of DBH-immunopositive neurons decreased by 32% 48 h after the introduction of anti-DBH–saporin into LC (Fig. 1).
Fig. 1. Dopamine-β-hydroxylase-immunopositive neurons in the locus coeruleus (LC) of neonatal rats 48 h after the introduction into the lateral ventricle of the brain of 0.9% NaCl in the control (a) and 0.5 µg of anti-DBH–saporin in the experiment (b); the number of dopamine-β-hydroxylase-immunopositive neurons in the left part of LC (c); n = 4, * p ≤ 0.05 (control compared to experiment). Symbols: IV – 4th ventricle of the brain. Scale bar: 100 µm.
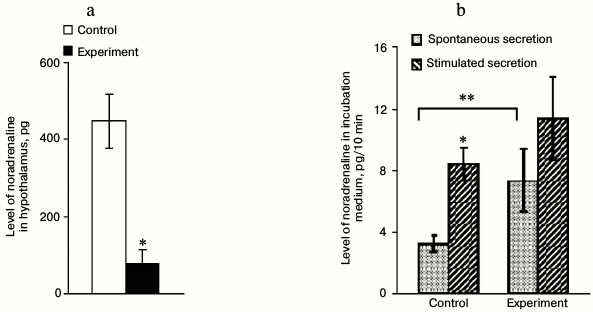
Noradrenaline content in and its secretion from hypothalamus after introduction of anti-DBH–saporin. The content of NA in the hypothalamus decreased 5.6-fold 72 h after the introduction of anti-DBH–saporin into rat brain on day 2 of life, being evidence of functional insufficiency of NA-synthesizing neurons of the brain in the given model (Fig. 2a). The NA level averaged for three 10-min fractions in the incubation medium (spontaneous secretion) increased 2-fold in the experiment compared to the control. The stimulated secretion in the control was >2-fold higher than the spontaneous secretion, while the experiment showed no reliable differences between these two parameters (Fig. 2b).
Fig. 2. Level of noradrenaline (NA) in hypothalamic tissue after incubation (a) and in the incubation medium (spontaneous and stimulated secretion) (b) expressed in pg per 10 min, after introduction into the lateral ventricle of the brain of 0.9% NaCl in the control and 0.5 µg of anti-DBH–saporin in the experiment; n = 6, (a) * p ≤ 0.05 (control compared to experiment); (b) * p ≤ 0.05 (spontaneous secretion compared to stimulated secretion), ** p ≤ 0.05 (control compared to experiment).
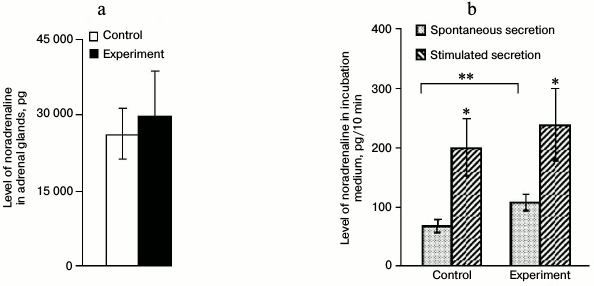
Noradrenaline content in and its secretion from adrenal glands after introduction of anti-DBH–saporin. The content of NA in adrenal tissues did not change 72 h after the introduction of anti-DBH–saporin into rat brain on day 2 of life compared to the control (Fig. 3a). The NA level in the incubation medium averaged for three 10-min fractions (spontaneous secretion) in the experiment was 1.6-fold higher compared to the control. The stimulated vs. spontaneous secretion was the same in the control and in the experiment (Fig. 3b).
Fig. 3. Level of noradrenaline (NA) in adrenal tissue after incubation (a) and in the incubation medium (spontaneous and stimulated secretion) (b), expressed in pg per 10 min, after introduction into the lateral ventricle of the brain of 0.9% NaCl in the control and 0.5 µg of anti-DBH–saporin in the experiment; n = 6, (b) * p ≤ 0.05 (spontaneous secretion compared to stimulated secretion), ** p ≤ 0.05 (control compared to experiment).
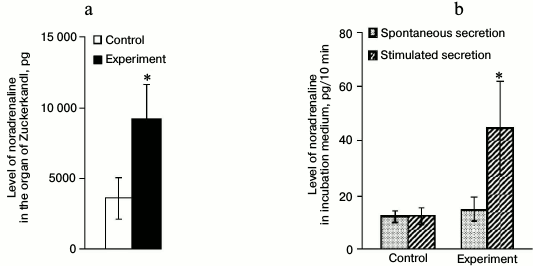
Noradrenaline content in and its secretion from OZ after introduction of anti-DBH–saporin. The content of NA in OZ tissues increased 2.5-fold 72 h after the introduction of anti-DBH–saporin into rat brain on day 2 of life after the incubation (Fig. 4a). The NA level in the incubation medium averaged for three 10-min fractions (spontaneous secretion) in the experiment did not change compared to the control. The stimulated secretion did not differ from the spontaneous secretion in the control but was 2.5-fold higher in the experiment (Fig. 4b).
Fig. 4. Level of noradrenaline (NA) in OZ after incubation (a) and in the incubation medium (spontaneous and stimulated secretion) (b), expressed in pg per 10 min, after introduction into the lateral ventricle of the brain of 0.9% NaCl in the control and 0.5 µg of anti-DBH–saporin in the experiment; n = 6, (a) * p ≤ 0.05 (control compared to experiment); (b) * p ≤ 0.05 (spontaneous secretion compared to stimulated secretion).
DISCUSSION
Understanding which NA sources exist in an organism in the critical period of morphogenesis is important for assessing the role of NA at that time. The secretion of NA from adrenal glands and extra adrenal chromaffin cells into the general circulatory system is undisputed [14], while the question whether NA is secreted from the brain into blood arose only after the development and confirmation of our hypothesis that the brain plays the role of a multipotent endocrine organ before BBB formation [22]. We have shown for the first time how the endocrine organ secretes NA into the general circulatory system of rats in the perinatal period of their development [10, 11]. Therefore, the main purpose of this work was to show the integration of the brain into the system of NA-secreting organs and its participation in their reciprocal humoral regulation. We have verified this hypothesis using a model of functional inhibition of NA production by the brain and its secretion into the general circulatory system by means of anti-DBH–saporin.
As it is known, NA is synthesized in the secretory granules through dopamine conversion into NA with by DBH, which is present in the granules in the free form or incorporated into the granule membrane. During the release of granule contents from the cells via exocytosis, the granule membrane with the embedded DBH becomes a component of the plasma membrane and a target for the antibody–toxin molecular complex. Later, this complex enters the cells via endocytosis and the toxin is released into the cytoplasm, causing ribosome destruction and cell death.
Anti-DBH–saporin introduced into the lateral ventricles of brain has a specific effect only on NA-ergic neurons [23]. Previously, when developing this model, we showed that impairment of NA production in the brain of newborn rats resulted in its decreased concentration in blood plasma after 48 h, with suppression of NA synthesis in the brain but without any changes in its content in peripheral organs. After 72 h in the same model, the secretory activity of NA-synthesizing peripheral organs undergoes compensatory changes resulting not only in normalization of the NA level in blood plasma, but even its increase compared to the control [21]. These data demonstrate the reciprocal humoral regulation of NA-producing organs via autoreceptors, on one hand, and the inertness of this system, on the other hand. As a result, NA homeostasis is provided (i.e. the maintenance of physiologically active concentration of NA in the general circulatory system).
In this work, we attempted to provide a better understanding of the mechanisms of plasticity of the NA-producing organs that maintain NA homeostasis. We assessed the spontaneous and K+-stimulated secretion of NA from the NA-producing organs, viz. the brain (hypothalamus), the adrenal glands, and OZ in the model of selective destruction of NA-ergic neurons of the brain described above. When reproducing this model in our work, we provided additional immunohistochemical control of its efficiency by assessing the number of DBH-containing neurons in LC and showed a decrease in the number of DBH-immunopositive neurons by 32% after introduction of the toxin. Thus, the adequacy of the model of specific destruction of NA-ergic neurons of the brain has been confirmed morphologically.
We have assessed the NA content and production not in the entire brain but in the hypothalamus, because this structure is abundantly innervated with NA-ergic axons [7]. According to our data, in spite of the drastic decrease in NA content in the hypothalamus as a result of destruction of the NA-ergic system of the brain, there was an increase in spontaneous secretion of NA from the hypothalamus, which may be associated with the compensatory intensification of NA secretion by surviving neurons and is a manifestation of the mechanism of neuroplasticity aimed at maintenance of the physiologically active concentration of NA in peripheral blood. At the same time, the absence of response to stimulation in this model is most likely accounted for by depletion of the reserve pool that releases catecholamines under conditions of long-term (minutes) stimulation [24].
The analysis of NA secretion in the adrenal glands has shown that spontaneous secretion of NA increases after introduction of anti-DBH–saporin into the brain, while the content of NA in adrenal tissue and the stimulated secretion do not change compared to the control. These data demonstrate the compensatory intensification of NA secretion from a labile vesicular pool in case of inhibition of its synthesis in the brain. At the same time, the synthesis of NA in chromaffin cells of the adrenal glands and the state of the stable pool are maintained at the control level.
In OZ, we observed a quite different (compared to adrenal glands) pattern of NA secretion after inhibition of its synthesis in the brain of neonatal animals. The higher NA content in OZ tissue compared to the control may be due to intensification of NA synthesis. At the same time, spontaneous secretion does not change, but the pattern of stimulated secretion is different: in the control, there is no response of OZ to K+ depolarization, i.e. the levels of spontaneous and stimulated excretion are similar, while the inhibition of NA synthesis in the brain results in emergence of the ability to respond to stimulation. OZ is the largest assembly of extra-adrenal chromaffin tissue; in rats, it reaches the maximum functional activity by day 3 and then undergoes involution by the end of the second week of life [13]. In previous study, we have shown that the response of OZ to depolarization in ontogeny is absent in rats on day 3 of life but is observed on day 15 in the period of involution [25], i.e. the death of chromaffin cells of OZ is accompanied by a compensatory increase in the secretion of NA in living cells. It can be supposed that the same compensatory reaction of OZ in the form of enhanced synthesis and stimulated secretion of NA is also observed in the model used in the present work.
We believe that the mechanism of compensatory intensification of NA secretion revealed in this work is as follows: NA synthesized in the developing brain is secreted into the general circulatory system, providing endocrine regulation of the development and function of peripheral target organs via specific autoreceptors. The expression of adrenoceptor genes is found in the nerve and chromaffin cells of rats already in the prenatal period of their development [16].
Thus, in the critical period of morphogenesis, the brain is integrated into the system of NA-secreting organs and participates in their reciprocal humoral regulation, which is manifested in the compensatory intensification of NA secretion in each of the NA sources studied in this work during the specific inhibition of its synthesis in the brain. As a result of such humoral interaction between the central and peripheral sources of NA, its physiologically active concentration in the general circulatory system is maintained.
Acknowledgements
This work was supported by the Russian Science Foundation (project No. 14-15-01122).
REFERENCES
1.Berger-Sweeney, J., and Hohmann, C. F. (1997)
Behavioral consequences of abnormal cortical development: insights into
developmental disabilities, Behav. Brain Res., 86,
121-142.
2.Kreider, M. L., Seidler, F. J., Cousins, M., Tate,
C. A., and Slotkin, T. A. (2004) Transiently overexpressed
α2-adrenoceptors and their control of DNA synthesis in the
developing brain, Dev. Brain Res., 152, 233-239.
3.Hildreth, V., Anderson, R. H., and Henderson, D. J.
(2009) Autonomic innervation of the developing heart: origins and
function, Clin. Anat., 22, 36-46.
4.Viemari, J. C., Bevengut, M., Burnet, H., Coulon,
P., Pequignot, J. M., Tiveron, M. C., and Hilaire, G. (2004) Phox2a
gene, A6 neurons, and noradrenaline are essential for development of
normal respiratory rhythm in mice, J. Neurosci., 24,
928-937.
5.Thomas, S. A., Matsumoto, A. M., and Palmiter, R.
D. (1995) Noradrenaline is essential for mouse fetal development,
Nature, 374, 643-646.
6.Tischler, A. S. (1989) The rat adrenal medulla,
Toxicol. Pathol., 17, 330-332.
7.Ugrumov, M. V. (1999) Mechanisms of Endocrine
Regulation [in Russian], Nauka, Moscow.
8.Huber, K., Kalcheim, C., and Unsicker, K. (2009)
The development of the chromaffin cell lineage from the neural crest,
Auton. Neurosci., 151, 10-16.
9.Kostrzewa, R. M. (2007) The blood-brain barrier for
catecholamines – revisited, Neurotox. Res., 11,
261-271.
10.Zubova, Yu. O., Bondarenko, N. S., Sapronova, A.
Ya., and Ugrumov, M. V. (2015) Noradrenaline secretion from the brain
into the general circulatory system during the ontogenesis in rats,
Neirokhimiya, 32, 116-122.
11.Zubova, Yu. O., Bondarenko, N. S., Sapronova, A.
Ya., and Ugrumov, M. V. (2015) Simulation of chronic selective shutdown
of noradrenaline synthesis in the brain of neonatal rats, Dokl.
Akad. Nauk, 461, 608-611.
12.Verhofstad, A. A. J., Hokfelt, T., Goldstein, M.,
Steinbusch, H. W. M., and Joosten, H. W. J. (1979) Appearance of
tyrosine hydroxylase, aromatic amino-acid decarboxylase, dopamine
β-hydroxylase and phenylethanolamine N-methyltransferase during
the ontogenesis of the adrenal medulla, Cell Tissue Res.,
200, 1-13.
13.Schober, A., Parlato, R., Huber, K., Kinscherf,
R., Hartleben, B., Huber, T. B., Schutz, G., and Unsicker, K. (2013)
Cell loss and autophagy in the extra adrenal chromaffin organ of
Zuckerkandl are regulated by glucocorticoid signaling, J.
Neuroendocrinol., 25, 34-47.
14.Goldstein, D. S., Eisenhofer, G., and Kopin, I.
J. (2003) Sources and significance of plasma levels of catechols and
their metabolites in humans, J. Pharm. Exp. Ther., 305,
800-811.
15.Thomas, G. B., Cummins, J. T., Smythe, G.,
Gleeson, R. M., Dow, R. C., Fink, G., and Clarke, I. J. (1989)
Concentrations of dopamine and noradrenaline in hypophysial portal
blood in the sheep and the rat, J. Endocrinol., 121,
141-147.
16.Fujinaga, M., and Scott, J. C. (1997) Gene
expression of catecholamine synthesizing enzymes and
β-adrenoceptor subtypes during rat embryogenesis, Neurosci.
Lett., 231, 108-112.
17.Coradazzi, M., Gulino, R., Garozzo, S., and
Leanza, G. (2010) Selective lesion of the developing central
noradrenergic system: short- and long-term effects and reinnervation by
noradrenergic-rich tissue grafts, J. Neurochem., 114,
761-771.
18.Wiley, R., and Kline, R. (2000) Neuronal
lesioning with axonally transported toxins, Neurosci. Methods,
103, 73-82.
19.Ashwell, K. W. S., and Paxinos, G. (2008)
Atlas of the Developing Rat Nervous System, Vol. 3, Elsevier
Academic Press, San Diego.
20.Khazipov, R., Zaynutdinova, D., Ogievetsky, E.,
Valeeva, G., Mitrukhina, O., Manent, J. B., and Represa, A. (2015)
Atlas of the postnatal rat brain in stereotaxic coordinates, Front.
Neuroanat., 9, 161.
21.Nikishina, Yu. O., Murtazina, A. R., Sapronova,
A. Ya., Melnikova, V. I., Bondarenko, N. S., and Ugryumov, M. V. (2016)
Reciprocal humeral regulation of endocrine sources of noradrenaline in
the perinatal period of development in rats, Ontogenez,
47, 287-295.
22.Ugrumov, M. V. (2010) Developing brain as an
endocrine organ: a paradoxical reality, Neurochem. Res.,
35, 837-850.
23.Wrenn, C. C., Picklo, M. J., Lappi, D. A.,
Robertson, D., and Wiley, R. G. (1996) Central noradrenergic lesioning
using anti-DBH–saporin: anatomical findings, Brain Res.,
740, 175-184.
24.Rizzoli, S. O., and Betz, W. J. (2005) Synaptic
vesicle pools, Nat. Rev. Neurosci., 6, 57-69.
25.Bondarenko, N. S., Murtazina, A. R.,
Dilmukhametova, L. K., Ikonopistseva, M. A., Volina, E. V., and
Ugrumov, M. V. (2016) Secretory activity of the brain and peripheral
organs: spontaneous and stimulated secretion of noradrenaline during
the ontogenesis in rats, Dokl. Akad. Nauk, 467, 1-4.