Electrogenesis in Plasma Membrane Fraction of Halotolerant Microalga Dunaliella maritima and Effects of N,N′-Dicyclohexylcarbodiimide
L. G. Popova1,a*, D. A. Matalin1, and Yu. V. Balnokin1
1Timiryazev Institute of Plant Physiology, Russian Academy of Sciences, 127276 Moscow, Russia* To whom correspondence should be addressed.
Received June 22, 2020; Revised June 22, 2020; Accepted June 26, 2020
The effects of N,N′-dicyclohexylcarbodiimide (DCCD), non-specific inhibitor of various transport systems functioning in biological membranes, on Na+-transporting P-type ATPase of the green halotolerant microalga Dunaliella maritima were studied in the experiments with vesicular plasma membranes isolated from the alga cells. The effects of DCCD on electrogenic/ion transport function of the enzyme and its ATP hydrolase activity were investigated. Electrogenic/ion transport function of the enzyme was recorded as a Na+-dependent generation of electric potential on the vesicle membranes with the help of the potential-sensitive probe oxonol VI. It was found that unlike many other ion-transporting ATPases, the Na+-ATPase of D. maritima is insensitive to DCCD. This agent did not inhibit either ATP hydrolysis catalyzed by this enzyme or its transport activity. At the same time DCCD affected the ability of the vesicle membranes to maintain electric potential generated by the D. maritima Na+-ATPase. The observed effects can be explained based on the assumption that DCCD interacts with the Na+/H+ antiporter in the plasma membrane of D. maritima.
KEY WORDS: Dunaliella maritima, Na+-ATPase, N,N′-dicyclohexylcarbodiimide, microalgae, membrane potential, plasma membraneDOI: 10.1134/S0006297920080088
Abbreviations: Δψ, transmembrane electric potential; CCCP, carbonyl cyanide m-chlorophenylhydrazone; DCCD, N,N′-dicyclohexylcarbodiimide; ETH157, sodium ionophore II (N,N′-dibenzyl-N,N′-diphenyl-1,2-phenylenedioxydiacetamide); PM, plasma membrane.
INTRODUCTION
Animal and plant cells differ in the way the plasma membrane is energized. In animal cells, the main role in energization of the cell membrane belongs to the active transport of Na+ and K+, carried out by the Na+,K+-ATPase. This enzyme maintains a non-equilibrium distribution of Na+ and K+ across the membrane and creates a negative potential on the cytoplasmic side of the membrane [1, 2]. The gradient of Na+ electrochemical potential created by the ATPase provides the driving force for the processes of passive diffusion of other ions, as well as the processes of secondary active co-transport of various substances through the membrane. In plant cells, transport processes via the plasma membrane are maintained by a proton gradient, which is produced by the electrogenic H+ pump, H+-ATPase [3]. It moves protons from the cell into the environment/apoplast using metabolic energy, and this process is accompanied by generation of a negative electric potential on the cytoplasmic side of the membrane.
However, electrogenic Na+ pumps were also found in the plasma membrane (PM) in some representatives of the plant kingdom – halotolerant green microalgae Tetraselmis viridis and Dunaliella maritima [4, 5]. The Na+-pumps from microalgae belong to the family of P-type ATPases, which also includes the PM proton pump of higher plants and the Na+,K+-ATPase of animal cells [6, 7]. It is interesting to note that the Na+-ATPases in the PM coexist with the H+-pumps – P-type ATPases in these microalgae as demonstrated in the experiments on vesicular plasma membranes isolated from the alga cells [5, 8]. These experimental data are in agreement with the data obtained in the in silico analysis of the sequenced genomes of green (Chlophyta) microalgae Ostreococcus tauri and Chlamydomonas reinhardtii, which also shows coexistence of the Na+- and H+-ATPases of P-type in these organisms [9].
Green microalgae of the genus Dunaliella are convenient model objects for studying physiology of plant tolerance to abiotic stresses. They are able to withstand extreme environmental conditions including high salinity [10]. These algae lack a large central vacuole and therefore only mechanisms localized in the plasma membrane are responsible for ionic homeostasis of the cytoplasm. In particular, an electrogenic Na+-transporting ATPase was found in the plasma membrane of the halotolerant microalgae Dunaliella maritima, which was apparently responsible for Na+-homeostasis in this alga cells [5]. This paper is a continuation of the study of properties of D. maritima Na+-ATPase.
No conservative motifs responsible for the Na+ ion binding has been identified in the primary structure of Na+-transporting membrane proteins, and the ability to transfer Na+ has been assigned to the features of the three-dimensional architecture of the proteins. The conserved transmembrane region is similar to the structure of crown ether, where Na+ plays the role of central coordinating ion [11, 12]. Individual amino acid residues at the corresponding positions in the protein are likely required to form a structure that can selectively bind Na+ ions. It was established using Na+,K+-ATPases from animal cells and Na+-specific bacterial F1F0-ATPases as examples that specificity of these enzymes with respect to transferred cations can be changed by single amino acid substitutions in the protein [13, 14]. The first step in identifying such amino acid residues could be the study of interaction of the protein with certain inhibitors with known mechanism of action. N,N′-dicyclohexylcarbodiimide (DCCD), a nonspecific inhibitor of various transport systems that function in biological membranes including ATPases of different types: F-type [15-17], V-type [18, 19], and P-type [20, 21] is an example of such inhibitor. The mechanism of suppression of protein transport activity by this inhibitor is well established. DCCD is a carboxyl-modifying agent that covalently binds to a specific amino acid residue (usually Glu or Asp) in the transmembrane region of the transport system at the site where the transferred cation should be bound, and thereby inhibits activity of the transport system [15, 19].
This paper presents the results of experiments investigating possible interactions of D. maritima Na+-ATPase with DCCD, namely, we studied the effects of DCCD on electrogenic/ion-transport function of this enzyme and its ATP-hydrolase activity. The experiments were carried out with a fraction of membrane vesicles enriched with plasma membrane (PM) isolated from D. maritima cells.
MATERIALS AND METHODS
Plant material and growing conditions. The studies were performed using green halotolerant microalga Dunaliella maritima Massjuk [22] as an object. The alga was cultured in 1-liter glass cylinders (average thickness of the algae suspension layer was 5 cm) in a medium of the following composition ( mM): NaCl – 500, NaNO3 – 10, KH2PO4 – 5, MgSO4 – 6, Ca(NO3)2 – 1.5, FeSO4 – 0.01, EDTA – 0.1, 1 ml/liter of a solution of microelements prepared according to Abdullaev and Semenenko [23]; pH was adjusted to 8.0 with NaOH. The culture was continuously bubbled with air containing 1.5% CO2. Illumination of 15,000 lux was provided with a cool white luminescent lamps using 14 : 10 h light : dark cycles.
Isolation of plasma membrane vesicles from D. maritima cells. Isolation of plasma membrane (PM) vesicles from microalgae cells was carried out according to the method described in [5]. The membrane fraction enriched with PM was obtained by differential centrifugation and centrifugation in a discontinuous (30/38%) sucrose gradient. In all experiments, including determination of ATP hydrolase activity of the enzyme, a freshly isolated PM fraction was used.
The obtained PM preparations comprised a mixture of inverted (inside-out, cytoplasmic side facing the external medium) and normally oriented (right-side-out) vesicles. In the experiments activity of only the inverted vesicles was observed in which active centers of ATPase were exposed to the external medium.
Detection of electrogenic activity of D. maritima Na+-ATPase. Electrogenic activity of the Na+-ATPase in isolated PM vesicles was detected as Na+- and ATP-dependent generation of electric potential difference across the vesicle membrane, as described in [5]. Generation of electric potential across the vesicle membrane was monitored through changes in differential absorption (A621-A582) of the potential-sensitive optical probe oxonol VI. Measurements were carried out with a dual wavelength spectrophotometer Hitachi 557 in a two-wavelength differential absorption mode (measurement wavelength λ2 = 621 nm, reference wavelength λ1 = 582 nm). A standard reaction medium (in a 2-ml spectrophotometric cuvette) was of the following composition: 0.4 M sucrose, 5 mM MgSO4, 1 mM EGTA, 20 mM Mes/BTP buffer (pH 7.5), 2 µM oxonol VI, about 100 µg of the membrane protein. Generation of electric potential across the vesicular membranes was initiated by adding ATP (to the final concentration of 2 mM; ATP was used in the form of Tris salt) and Na+ ions (in the form of Na2SO4).
Analytical methods. ATP-hydrolase activity of the membrane fractions was determined from the amount of inorganic phosphate (Pi) released from the added ATP. ATP hydrolysis by the PM fractions was measured in the reaction mixture of the following composition: 0.4 M sucrose, 20 mM Mes/BTP buffer (pH 5.5-9.0), 1 mM MgSO4, 1 mM EGTA, 50 µM carbonyl cyanide m-chlorophenylhydrazone (CCCP), 10-15 µg of membrane protein. The reaction was initiated by adding 0.5 mM Na2ATP and stopped after 20 min. Other additives are indicated in the figure captions. The released Pi was tested using the Carter and Karl method [24].
Protein content in the PM preparations was determined by the micromethod of Simpson and Sonne [25]. The method is based on disintegration of membranes in an alkaline medium and determination of the intensity of the protein-Coomassie G-250 complex staining.
Fresh DCCD solutions prepared in ethanol were used in the work. Final ethanol concentration introduced with DCCD to the reaction mixtures during measurements of ATPase activity did not exceed 0.2%. Control samples contained same concentrations of ethanol.
Figures and graphs show representative experimental data from at least three independent replicates.
RESULTS AND DISCUSSION
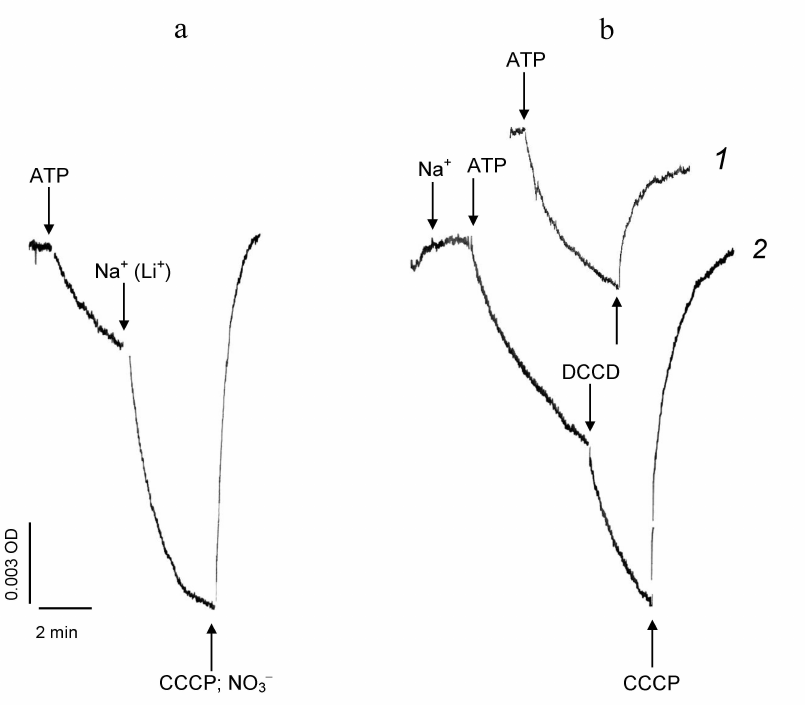
The electrogenic Na+-transporting ATPase operates in the plasma membrane of the microalgae D. maritima, as has been shown earlier in the experiments with PM vesicles isolated from the alga cells [5]. Functions of the D. maritima electrogenic Na+-ATPase is illustrated in Fig. 1a. ATP addition to the PM vesicle suspension induces generation of electric potential (Δψ) across the vesicle membranes due to operation of D. maritima H+-ATPase [5]. Sodium addition leads to a significant stimulation of Δψ generation across the vesicle membranes, which is obviously due to the transfer of Na+ ions to the vesicle lumen carried out by the Na+-ATPase. Generation of Δψ across the vesicle membranes can be also induced by Li+, but not by K+ (not shown). Such ionic specificity is typical for many membrane Na+-transport mechanisms, which can transport Li+ along with Na+ ions [26, 27].
DCCD addition to the reaction mixture in the absence of Na+ ions during the ATP-dependent generation of Δψ across the vesicle membranes caused decay of the formed potential (Fig. 1b, curve 1). However, when DCCD was added to the suspension of PM vesicles during ATP- and Na+-dependent generation of Δψ, the rate of its generation did not decrease (which would be expected in the case of Na+-dependent ATPase inhibition), but even increased (Fig. 1b, curve 2).
Fig. 1. ATP- and Na+-dependent generation of electric potential across the plasma membrane vesicles from D. maritima. Effects of DCCD. Generation of electric potential (Δψ) across the vesicle membranes (“+” in the vesicle lumen) is manifested by the decrease in the differential absorption of the potential-sensitive probe oxonol VI. Generation of Δψ was initiated by addition of ATP (in the form of Tris salt) to a final concentration of 1 mM. The time points of additive introductions are indicated by arrows. a) Initial: standard reaction mixture without Na+. Addition of Na+ (in the form of 10 mM Na2SO4) accelerates Δψ generation (a similar effect is observed with addition of Li+). The protonophore CCCP (50 µM) or permeant anion NO3– (50 mM, in the form of BTP NO3 salt, pH 7.5) reverses differential absorption of the probe to the initial level, indicating decay of the formed Δψ (due to the exit of H+ from the vesicle lumen or due to the compensation of the excess positive charge inside the vesicles by the anion, respectively). b) Effects of DCCD (100 µM) added to the reaction mixture during Δψ generation across the vesicle membranes. Curve 1: standard reaction medium without Na+. Curve 2: standard reaction medium containing Na+ (as 1 mM Na2SO4).
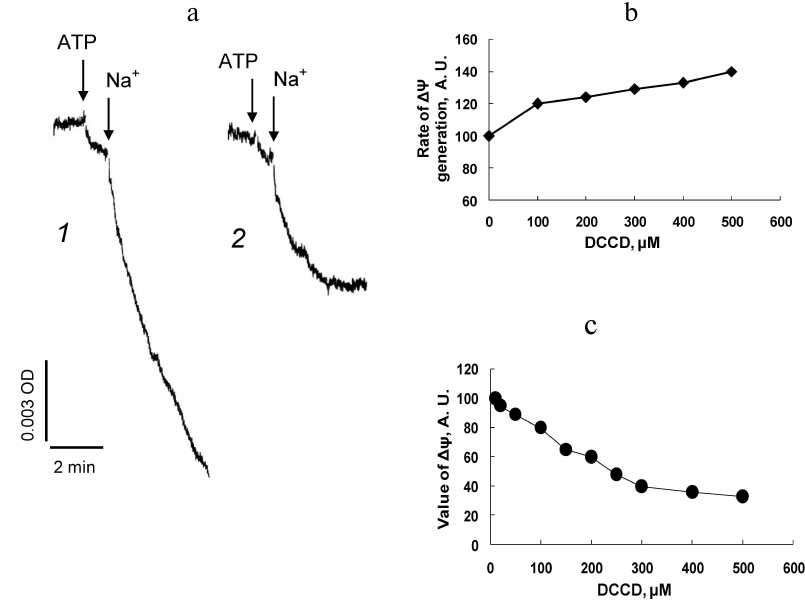
It is known that the transported ion protects transport system from inhibition by DCCD [16, 17, 28]. In order to prevent the possible protective effect of Na+ ions on the Na+-ATPase, PM vesicles were pre-incubated with DCCD for 15 min in the absence of Na+, after which generation of Δψ across the vesicle membrane was induced by sequential addition of ATP and Na+. In this case the initial rate of ATP- and Na+-induced generation of Δψ also increased, depending on concentration of DCCD present in the reaction mixture (Fig. 2, a and b), but the value of Δψ across the vesicle membrane generated by the Na+-ATPase decreased (Fig. 2, a and c). Increase in the initial rate of Δψ generation indicates the absence of DCCD inhibitory effect on Na+-ATPase. This was confirmed by the results of experiments examining ATP-hydrolase activity of this enzyme.
Fig. 2. Effects of DCCD on electric potential generated by Na+-ATPase across the plasma membrane vesicles from D. maritima. The vesicles were incubated (15 min, 22°C) in the standard reaction mixture containing DCCD, after which Δψ generation was initiated by sequential addition of ATP and Na+ (as 10 mM Na2SO4). a) Kinetics of Δψ generation in the absence of DCCD (curve 1) and in the presence of 300 µM of DCCD (curve 2). Dependence of the initial rate of Δψ generation (b) and the stationary value of the generated Δψ (c) on DCCD concentration in the reaction mixture.
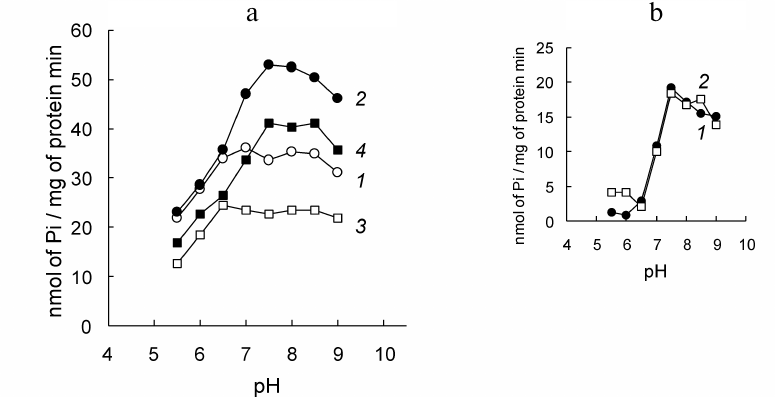
The PM fractions from D. maritima catalyze ATP hydrolysis in a wide pH range (Fig. 3). The Na+-stimulated increase in ATP hydrolysis reflects the ATP-hydrolase activity of Na+-ATPase, maximum of which is observed at pH 7.5-8.0 (Fig. 3a, curve 2). Pre-incubation of the PM vesicles with DCCD prior to addition of ATP and Na+ does not cause significant changes in the value of the Na+-stimulated increase in ATP hydrolysis (Fig. 3a, compare curves 1 and 2 in pairs, curves 3 and 4; Fig. 3b). The latter obviously indicates absence of the inhibition of Na+-ATPase by this agent.
Fig. 3. pH-profiles of ATP hydrolysis catalyzed by the plasma membrane (PM) fractions from D. maritima and effects of DCCD. a) Standard reaction mixture contained additionally. Curves: 1) 25 mM KNO3; 2) 25 mM NaNO3; 3) 25 mM KNO3, 100 µM DCCD; 4) 25 mM NaNO3, 100 µM DCCD. In the samples with DCCD, the vesicles were pre-incubated in the standard reaction mixture containing additionally DCCD for 15 min prior to addition of Na+ and ATP. b) Na+-induced increase in ATP hydrolysis catalyzed by the PM fractions in the absence (curve 1) and in the presence of DCCD (curve 2).
The decrease in the stationary value of Δψ generated by the Na+-ATPase across the membrane of the vesicles pre-incubated with DCCD can be explained by the increase in ionic conductivity of the membrane resulted from interaction of DCCD with some ion-transporting protein in the PM from D. maritima other than the Na+-ATPase. Possible candidates for this role are H+-ATPase and/or Na+/H+ antiporter, which are also present in this membrane [5, 29]. Both H+-ATPase and Na+/H+ antiporter can interact with DCCD [30-32].
The assumption that pre-incubation of PM vesicles with DCCD leads to the increase of ionic conductivity of the membrane was verified in the experiments, where electric potential across the vesicle membrane was created in an ATP-independent manner, in the form of a Na+- or K+-diffusion potential, by introducing a cation and the corresponding ionophore, sodium ionophore II (N,N′-dibenzyl-N,N′-diphenyl-1,2-phenylenedioxydiacetamide) (ETH157) for Na+ and valinomycin for K+, to the vesicle suspension (Fig. 4).
Fig. 4. Effects of DCCD on the Na+-diffusion potential and K+-diffusion potential generated across the plasma membrane (PM) vesicles from D. maritima. Generation of Δψ across the vesicle membranes (“+” in the vesicle lumen) was initiated by introducing 10 mM Na2SO4 + 3 µM ETH-157 (a, b) or 10 mM K2SO4 + 3 µM valinomycin (c, d). DCCD concentration in the reaction mixture was 100 µM. a) DCCD was added after formation of the Na+- diffusion potential. b-d) PM vesicles were pre-incubated for 15 min in the standard reaction mixture containing 100 µM DCCD before the addition of remaining additives (10 mM Na2SO4 + 3 µm ETH-157 or 10 mM K2SO4 + 3 µM valinomycin). CCCP protonophore (50 µM) (a, c) reverses differential absorption of oxonol VI to the initial level, which indicates decay of the formed Δψ. d) Addition of 10 mM Na2SO4 leads to the decay of the formed Δψ.
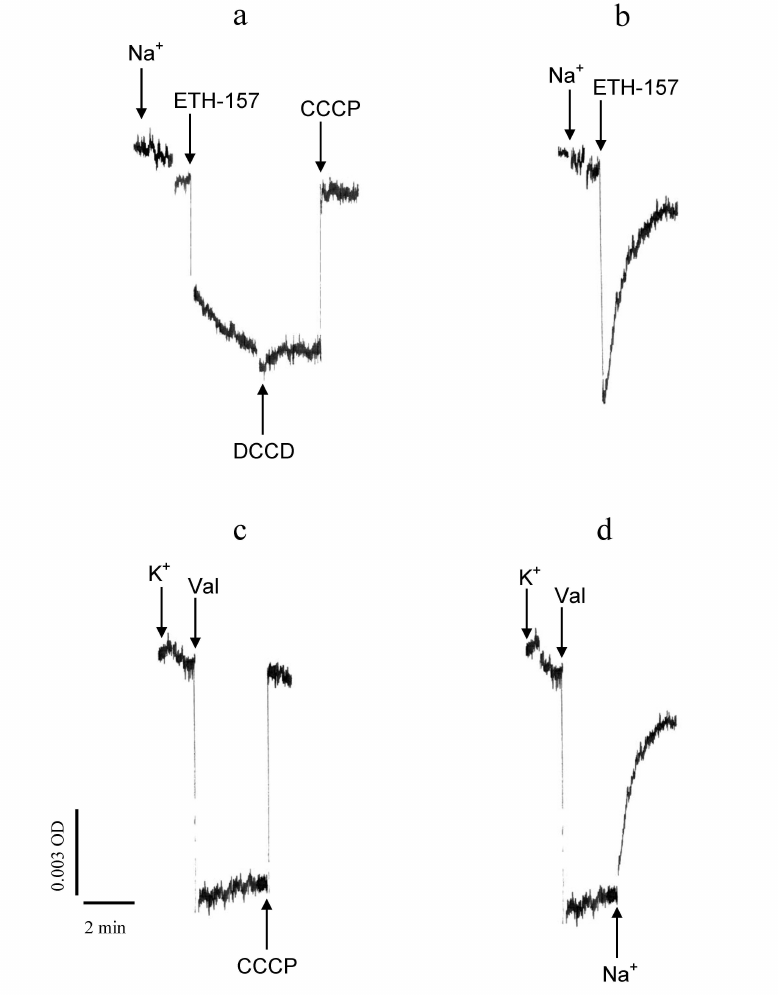
DCCD added to the vesicles did not produce any visible effect on the Na+-diffusion potential created across their membranes prior to that (Fig. 4a). However, if the vesicles were pre-incubated with DCCD before Na+ addition, the diffusion potential formed across the vesicle membranes upon Na+ application rapidly and spontaneously decayed (Fig. 4b). In the case when K+-diffusion potential was generated across the vesicle membranes pre-incubated with DCCD, it was maintained for a long period of time and decayed only when the protonophore CCCP was introduced to the reaction mixture (Fig. 4c). At the same time, the K+-diffusion potential generated across the vesicle membranes pre-incubated with DCCD decayed upon Na+ addition (Fig. 4d). These results allow concluding that dissipation of the electric potential across the vesicle membranes pre-incubated with DCCD depends on the presence of Na+ in the reaction mixture.
It should be noted that in the described experiments the transmembrane electric potential decayed due to the exit of cations other than Na+ (or accordingly K+) from the vesicle lumen. Since, in the presence of ETH157 (respectively, valinomycin), that provides high conductivity of the vesicle membrane for Na+ (or K+), the latter is in the state of electrochemical equilibrium on both sides of the vesicle membrane (“+” inside the vesicles) and, therefore, there are no driving forces for the exit of Na+ (or K+) from the vesicles. It is most likely that in this case the excess of positive charge from the vesicle lumen is carried away by a proton, and Na+-diffusion (or K+-diffusion) potential serves as a driving force for its exit. Obviously, the path of H+ leakage is formed due to interaction of DCCD with some protein(s) residing in the vesicular membrane. This path is potential-dependent, since the Na+-diffusion potential could be created across the vesicle membranes pre-incubated with DCCD, and the decay of the potential begins only when the certain potential value is reached (Fig. 4b).
Thus, the obtained data indicate that: (i) D. maritima Na+-ATPase is insensitive to DCCD, (ii) in the absence of Na+ ions, DCCD interacts with some protein/proteins of the vesicle membranes, (iii) Na+ protects this protein from interaction with DCCD, (iv) interaction of DCCD with this protein leads to the increase in H+ conductivity of the vesicle membrane, and the latter requires the presence of Na+ ions, (v) transmembrane electric potential is necessary for the DCCD-induced Na+-dependent H+-conductivity of the membrane to be realized.
The most obvious assumption is that the observed effects arise from the interaction of DCCD with the Na+/H+ antiporter in the PM of D. maritima. The antiporter is able to bind and transfer both Na+ and H+ through the membrane. Interaction with DCCD results in preserving ability of the antiporter to transfer H+ but not Na+. However, the presence of Na+ ion, probably bound at a specific protein site, is necessary for H+ transfer to occur. In other words, the electrically neutral cation-exchanging Na+/H+ antiporter is transformed into a potential-sensitive H+ uniporter when interacting with DCCD.
The question remains why the initial rate of ATP- and Na+-dependent potential generation across the membranes of the vesicles pre-incubated with DCCD increases (Fig. 2b), while DCCD does not stimulate ATP-hydrolase activity of the Na+-ATPase (Fig. 3).
Possible explanation for this phenomenon may be the following. Apparently, the Na+/H+ antiporter is not the only protein in D. maritima PM that DCCD binds to. DCCD can bind to the H+ pump operating in this membrane. DCCD is known to inactivate H+-ATPase blocking proton translocation by this enzyme [15, 19, 30]. Thus, interaction of DCCD with the H+-ATPase (and probably with certain ion channels also) leads to the decrease in total proton conductivity of the membrane, which, in turn, under experimental conditions, leads to the decrease in ion (proton) leaks from the vesicle lumen and increase of the electric potential generated across the vesicle membranes when the Na+-ATPase is turned on. As the certain values of Δψ are reached across the vesicle membranes during the Na+-ATPase operation, the proton conductivity of the membranes increases sharply due to increase in the proton conductivity of the Na+/H+ antiporter, which presumably is transformed as a result of interaction with DCCD from the electroneutral cation-exchange protein into the potential-sensitive H+-uniporter.
A primary Na+ pump, an electrogenic Na+ transporting P-type ATPase operates in the plasma membrane of the halotolerant microalgae D. maritima (Phylum Chlorophyta, class Chlorophyceae), which is responsible for Na+ homeostasis in this organism [5]. It was demonstrated that this enzyme was insensitive to DCCD unlike many other ion-transporting ATPases. DCCD did not inhibit either ATP hydrolysis catalyzed by this enzyme or its transport activity. Na+-ATPase of D. maritima was similar to the Na+-ATPase of another halotolerant microalga, Tetraselmis viridis (Chlorophyta, Prasinophyceae) in its insensitivity to DCCD [4]. Previously, it was also shown for the T. viridis Na+-ATPase that DCCD did not inhibit this enzyme [8]. Insensitivity of the Na+-transporting ATPases from two alga species to DCCD may reflect structural similarity of these enzymes emerging from their evolutionary proximity.
Funding. This work was financially supported by the Russian Foundation for Basic Research (project No. 20-04-00903).
Ethics declarations. The authors declare no conflict of interests in financial or any other sphere. This article does not contain any studies with human participants or animals performed by any of the authors.
REFERENCES
1.Boldyrev, A. A. (2001) Na/K-ATPase as an oligomeric
ensemble, Biochemistry (Moscow), 66, 821-831,
doi: 10.1023/a:1011964832767.
2.Scheiner-Bobis, G. (2002) The sodium pump, Eur.
J. Biochem., 269, 2424-2433,
doi: 10.1046/j.1432-1033.2002.02909.x.
3.Gaxiola, R. A., Palmgren, M. G., and Schumacher, K.
(2007) Plant proton pumps, FEBS Lett., 581, 2204-2214,
doi: 10.1016/j.febslet.2007.03.050.
4.Balnokin, Y. V., and Popova, L. G. (1994) The
ATP-driven Na+-pump in the plasma membrane of the marine
unicellular alga Platymonas viridis, FEBS Lett.,
343, 61-64, doi: 10.1016/0014-5793(94)80607-1.
5.Popova, L. G., Shumkova, G. A., Andreev, I. M., and
Balnokin, Y. V. (2005) Functional identification of electrogenic
Na+-translocating ATPase in the plasma membrane of the
halotolerant microalga Dunaliella maritima, FEBS Lett.,
579, 5002-5006, doi: 10.1016/j.febslet.2005.07.087.
6.Axelsen, K., and Palmgren, M. G. (1998) Evolution
and substrate specificities in the P-type ATPase superfamily, J.
Mol. Evol., 46, 84-101, doi: 10.1007/pl00006286.
7.Palmgren, M. G., and Nissen, P. (2011) P-type
ATPases, Annu. Rev. Biophys., 40, 243-266,
doi: 10.1146/annurev.biophys.093008.131331.
8.Pagis, L. Y., Popova, L. G., Andreev, I. M., and
Balnokin, Y. V. (2003) Comparative characterization of the two primary
pumps, H+-ATPase and Na+-ATPase, in the plasma
membrane of the marine alga Tetraselmis viridis, Physiol.
Plant., 118, 514-522,
doi: 10.1034/j.1399-3054.2003.00113.x.
9.Pedersen, Ch. N. S., Axelsen, K. B., Harper, J. F.,
and Palmgren, M. G. (2012) Evolution of plant P-type ATPases, Front.
Plant Sci., 3, doi: 10.3389/fpls.2012.00031.
10.Oren, A. (2005) A hundred years of
Dunaliella research: 1905-2005, Saline Systems, 1,
1-14, doi: 10.1186/1746-1448-1-2.
11.Gouaux, E., and MacKinnon, R. (2005) Principles
of selective ion transport in channels and pumps, Science,
310, 1461-1465, doi: 10.1126/science.1113666.
12.Meier, T., Krah, A., Bond, P. J., Pogoyelov, D.,
Diederichs, K., and Faraldo-Gomez, J. D. (2009) Complete
ion-coordination structure in the rotor ring of
Na+-dependent F-ATP synthases, J. Mol. Biol.,
391, 498-507, doi: 10.1016/j.jmb.2009.05.082.
13.Imagawa, T., Yamamoto, T., Kaya, Sh., Sakaguchi,
K., and Taniguchi, K. (2005) Thr-774 (transmembrane segment M5),
Val-920 (M8), and Glu-923 (M9) are involved in
Na+-transport, and Gln-923 (M8) is essential for
Na+,K+-ATPase activity, J. Biol. Chem.,
280, 18736-18744, doi: 10.1074/jbc.M500137200.
14.Kaim, G., and Dimroth, P. (1994) Construction,
expression and characterization of a plasmid-encoded
Na+-specific ATPase hybrid consisting of Propionigenium
modestum Fo-ATPase and Escherichia coli
F1-ATPase, Eur. J. Biochem., 222, 615-623,
doi: 10.1111/j.1432-1033.1994.tb18904.x.
15.Toei, M., and Noji, H. (2013) Single-molecule
analysis of F0F1-ATP synthase inhibited by
N,N′-dicyclohexylcarbodiimide, J. Biol. Chem., 288,
25717-25726, doi: 10.1074/jbc.M113.482455.
16.Ferguson, S., Keis, S., and Cook, G. M. (2006)
Biochemical and molecular characterization of a
Na+-translocating F1F0-ATPase from the
thermoalkaliphilic bacterium Clostridium paradoxum, J.
Bacteriol., 188, 5045-5054, doi: 10.1128/JB.
00128-06.
17.Soontharapirakkul, K., and Incharoensakdi, A.
(2010) Na+-stimulated ATPase of alkaliphilic halotolerant
cyanobacterium Aphanothece halophytica translocates
Na+ into proteoliposomes via Na+ uniport
mechamism, BMC Biochemistry, 11,
doi: 10.1186/1471-2091-11-30.
18.Murata, T., Kawano, M., Igarashi, K., Yamato, I.,
and Kakinuma, Y. (2001) Catalytic properties of
Na+-translocating V-ATPase in Enterococcus
hirae, Biochim. Biophys. Acta, 1505, 75-81,
doi: 10.1016/S0005-2728(00)00278-4.
19.Yokoyama, K., Nakano, M., Imamura, H., Yoshida,
M., and Tamakoshi, M. (2003) Rotation of the proteolipid ring in
the V-ATPase, J. Biol. Chem., 278,
24255-24258, doi: 10.1074/jbc.M303104200.
20.Corbalan-Garsia, S., Teruel, J. A., and
Gomez-Fernandez, J. C. (1992) Characterization of Ruthenium Red-binding
sites of the Ca2+-ATPase from sarcoplasmic reticulum and
their interaction with Ca2+-binding sites,
Biochemistry, 287, 767-774,
doi: 10.1042/bj2870767.
21.Wiangnon, K., Raksajit, W., and Incharoensakdi,
A. (2007) Presence of a Na+-stimulated P-type ATPase in the
plasma membrane of the alkaliphilic halotolerant cyanobacterium
Aphanothece halophytica, FEMS Microbiol. Lett.,
270, 139-145, doi: 10.1111/j.1574-6968.2007.00667.x.
22.Massyuk, N. P. (1973) Morphology, taxonomy,
ecology and geographic distribution of the genus Dunaliella Teod. and
prospects for its potential utilization, Naukova Dumka, Kiev, p.
244.
23.Abdullaev, A. A., and Semenenko, V. E. (1974)
Intensive cultivation and certain physiological characteristics of
Dunaliella salina Teod., Soviet Plant Physiol.,
21, 947-955.
24.Carter, S. G., and Karl, D. W. (1982) Inorganic
phosphate assay with malachite green: an improvement and evaluation,
J. Biochem. Biophys. Methods, 7, 7-13,
doi: 10.1016/0165-022x(82)90031-8.
25.Simpson, I. A., and Sonne, O. (1982) A simple,
rapid and sensitive method for measuring protein concentration in
subcellular membrane fractions prepared by sucrose density
ultracentrifugation, Anal. Biochem., 119, 424-427,
doi: 10.1016/0003-2697(82)90608-x.
26.Krulwich, T. A. (1983)
Na+/H+ antiporters, Biochim. Biophys.
Acta, 726, 245-264,
doi: 10.1016/0304-4173(83)90011-3.
27.Pagis, L. Ya., Popova, L. G., Andreev, I. M., and
Balnokin, Yu. V. (2001) Ion specificity of Na+-transporting
systems in the plasma membrane of the halotolerant alga Tetraselmis
(Platymonas) viridis, Russ. J. Plant Physiol., 48,
281-286, doi: 10.1023/A:1016645829002.
28.Kluge, C., and Dimroth, P. (1993) Specific
protection by Na+ or Li+ of the
F1Fo-ATPase of Propionigenium modestum
from the reaction with dicyclohexylcarbodiimide, J. Biol. Chem.,
268, 14557-14560.
29.Popova, L. G., Shumkova, G. A., Andreev, I. M.,
and Balnokin, Yu. V. (2000) Na+-dependent electrogenic
ATPase from the plasma membrane of the halotolerant microalga
Dunaliella maritima, Doklady Biochemistry, 375,
235-238, doi: 10.1023/A:1026675923730.
30.Sussman, M. R., and Slayman, C. W. (1983)
Modification of the Neurospora crassa plasma membrane
H+-ATPase with N,N′-dicyclohexylcarbodiimide, J.
Biol. Chem., 258, 1839-1843.
31.Kinsella, J. L., Wehrle, J., Wilkins, N., and
Sacktor, B. (1987) Inhibition of Na+-H+-exchange
by N,N′-dicyclohexylcarbodiimide in isolated rat renal brush
border membrane vesicles, J. Biol. Chem., 262,
7092-7097.
32.Murakami, N., and Konishi, T. (1989) Mechanism of
function of dicyclohexylcarbodiimide-sensitive
Na+/H+-antiporter in Halobacterium
halobium: pH effect, Arch. Biochem. Biophys., 271,
515-523, doi: 10.1016/0003-9861(89)90303-2.