HYPOTHESIS: Receptor Regulation of Senile Phenoptosis
M. V. Skulachev1,2, F. F. Severin3,4, and V. P. Skulachev2,3,4*
1Biological Faculty, Lomonosov Moscow State University, 119991 Moscow, Russia2Institute of Mitoengineering, Lomonosov Moscow State University, 119991 Moscow, Russia
3Belozersky Institute of Physico-Chemical Biology, Lomonosov Moscow State University, 119991 Moscow, Russia; E-mail: skulach@belozersky.msu.ru
4Department of Bioengineering and Bioinformatics, Lomonosov Moscow State University, 119991 Moscow, Russia; E-mail: skulach@genebee.msu.ru
* To whom correspondence should be addressed.
Received June 20, 2014
Here we present a concept that considers organism aging as an additional facultative function promoting evolution, but counterproductive for an individual. We hypothesize that aging can be inhibited or even arrested when full mobilization of all resources is needed for the survival of an individual. We believe that the organism makes such a decision based on the analysis of signals of special receptors that monitor a number of parameters of the internal and external environment. The amount of available food is one of these parameters. Food restriction is perceived by the organism as a signal of coming starvation; in response to it, the organism inhibits its counterproductive programs, in particular, aging. We hypothesize that the level of protein obtained with food is estimated based on blood concentration of one of the essential amino acids (methionine), of carbohydrates – via glucose level, and fats – based on the level of one of the free fatty acids. When the amount of available food is sufficient, these receptors transmit the signal allowing aging. In case of lack of food, this signal is cancelled, and as a result aging is inhibited, i.e. age-related weakening of physiological functions is inhibited, and lifespan increases (the well-known geroprotective effect of partial food restriction). In Caenorhabditis elegans, lowering of the ambient temperature has a similar effect. This geroprotective effect is removed by the knockout of one of the cold receptors, and replacement of the C. elegans receptor by a similar human receptor restores the ability of low temperature to increase the lifespan of the nematode. A chain of events linking the receptor with the aging mechanism has been discovered in mice – for one of the pain receptors in neurons, the nerve endings of which entwine pancreas β-cells. Age-related activation of these receptors inhibits the work of insulin genes in β-cells. Problems with insulin secretion lead to oxidative stress, chronic inflammation, and type II diabetes, which can be regarded as one of the forms of senile phenoptosis. In conclusion, we consider the role of some psychological factors in the regulation of the aging program.
KEY WORDS: phenoptosis, aging, aging program regulation, food restriction, diabetesDOI: 10.1134/S0006297914100022
Abbreviations: mROS, mitochondrial reactive oxygen species; ROS, reactive oxygen species; SkQ, derivatives of plastoquinone and penetrating cations (Sk+); SkQ1, plastoquinonyl decyltriphenylphosphonium.
One of the predictions of the hypothesis of programmed aging is that
aging can be controlled by regulatory systems of the organism [1, 2]. With this concept, aging is
nothing but a phenomenon counterproductive for an individual, but
useful for its evolution. If this is true, it is logical for an
organism to be able to slow or even cancel aging in response to a sharp
deterioration in living conditions, when the very survival of an
individual is threatened [2]. If aging is caused by
slow self-poisoning of our cells by reactive oxygen species (ROS) [1-3], termination of this
self-poisoning might prove to be the reserve that would save the
individual at a critical moment [3]. Such a concept
implies the presence of specific receptors that perceive the signal of
the critical situation and transfer it to the hypothetical
“center of aging management”. If the signal is strong
enough and stable over time, the “center” will send an
order to reduce ROS level, i.e. to inhibit aging. The inhibition can be
removed later, when the situation becomes once again normal and the
individual will be able to take care of not only its own survival, but
also of acceleration of evolution of its offspring.
FOOD RESTRICTION AS A SIGNAL INHIBITING THE AGING PROGRAM
Food restriction is an example of one of the most universal ways to slow the aging program due to signals of certain receptors. From 1934, K. M. MacCay and T. B. Robertson published a series of articles on the 50-70% increase in lifespan of mice and rats restricted in food consumption [4-7]. It is also worth noting that the cases of death caused by lung infections and some tumors dramatically decreased in these animals. They observed that those long-living animals looked young and agile during their entire lifetime regardless of age. Later, the beneficial effect of food restriction on lifespan was demonstrated for a wide variety of eukaryotic species ranging from yeast to rhesus macaques and humans. Harman’s hypothesis on the role of ROS in aging [8] led to a new explanation for this phenomenon: the most likely cause was seen to be the reduction of the amount of food oxidized by oxygen and hence the decrease in ROS production and their damaging effect on biopolymers. However, experiments have shown that it is not that simple. Early studies conducted in the same group of MacCay showed that heat production per 1 kg of body weight was even somewhat higher in the experimental cohort of rats restricted in food consumption than in the control [9].
Later, it became clear that it is enough for young Drosophila flies to starve for 7 or even only 2 days to turn into a long-living individual; the effect was similar when the flies were restricted in food consumption for its entire life [10, 11]. In addition, it was found that not only the increased amount of food, but also its smell reduced the geroprotective effect of food restriction in Drosophila and the nematode Caenorhabditis elegans [12, 13]. All these observations are not characteristic only for invertebrates. Already in 1934, Robertson et al. discovered that it was sufficient for mice to starve for two days per week to prolong their lifespan by 50-60% [5]. Moreover, it was shown that rats, mice and golden hamsters restricted in food by 50% for 2 years lived 20% longer than the control group, and food restriction for 1 year caused 40-60% increase in lifespan [14].
Further research has shown that carbohydrate, fat, and protein components contribute to the effect of food restriction. In relation to proteins, it was found that a single amino acid, methionine, is responsible for this effect [15-19]. Methionine one of the essential amino acids since it cannot be synthesized in mammals, and its supply entirely depends on consumed food. It was found that an increase in lifespan, reduction of ROS generation in mitochondria, and decrease in oxidative damage of mitochondrial DNA could be achieved by using a diet with protein replaced by an isocaloric mixture of amino acids without methionine [17-20]. It is important that food restriction had no effect on the oxidation of nuclear DNA [21]. Recent observations by G. Barja et al. are quite demonstrative [19]. According to these authors, a methionine-free diet for 7 weeks reduced ROS generation by liver mitochondria of old rats and decreased or even completely reversed oxidative damage to mitochondrial DNA, proteins, lipids, and carbohydrates in aging animals.
We think that the organism perceives food restriction as an alarming signal of impending starvation that could question the very existence of the population. To prevent this, the organism tries to slow (or even cancel) the aging program, which deliberately weakens the individual, so as to increase the pressure of natural selection [22, 23]. (In this context, it is interesting to mention that food restriction has no geroprotective effect on the queen bee, whose aging program is apparently canceled [24].) Food restriction is known to reduce oxidative stress [25-28]. In the framework of our concept, the increasing lifespan in this case is a result of a purely regulatory effect. For this reason such clearly signal effects as short fasting (or, to the contrary, the smell of food), and not only lifelong restriction or excess of food, deeply affect life cycle parameters. It is no coincidence that temporary food restriction (fasting) is better than constant fasting. A relatively short period of time is sufficient to transmit the signal, while prolonged starvation can exhaust the organism.
The signaling nature of the food restriction effect provides a good explanation to the experiments with methionine. Apparently, the organism measures the amount of consumed food, and above all essential amino acids required for biosynthesis of its proteins, by tracking the amount of only one amino acid, methionine, using some special receptor. Such receptors do exist for glucose, the level of which could be used by the organism to track the amount of carbohydrates in food. Analysis of the contribution of glucose decrease to the effect of food restriction is complicated by the fact that glucose metabolites can be formed not only of this monosaccharide, but also of a number of other sugars. There is no such complication in the case of methionine as a measure of the amount of protein component in food. It is quite possible that religious fasts are a way to prolong life by short periods of food restriction. People observing religious rituals are generally known to live longer [29].
It is logical to assume that regulation of aging by the fat component of food is similarly organized. In this case, the organism is likely to measure the level of some essential fatty acid (they belong to the group of unsaturated fatty acids).
It is important that food restriction not only increases average lifespan, but also prolongs youth, as observed by the discoverer of this phenomenon MacCay. Senile diseases recede, and the behavior and even the appearance of old animals often become indistinguishable from those of young individuals. In this regard, a recently published article by R. Weindruch et al. is quite demonstrative [30, 31]. They describe a 20-year-long experiment on 76 macaques that has shown that long-term 30% food restriction (adult animals aged 7-14 years were taken for the experiment) leads to a sharp decrease in age-dependent mortality (during 30 years only 20% of the “fasting” monkeys died, while the same parameter in the control group of animals receiving food ad libitum was 50%); disappearance of diabetes from the causes of death; 2-fold decrease in mortality from cancer (in macaques it is mainly intestinal adenocarcinoma); decrease in mortality from cardiovascular diseases; reduction of osteoporosis; prevention of the development of such senile signs as sarcopenia, reduction in the weight of brain gray matter, balding, graying, etc.
By the age of 30, 80% of the surviving control macaques demonstrated some signs of aging, while in the experimental group they were observed only in 20% of the animals. Figure 1 shows pictures of two monkeys of the same age (about 28 years). The control animal is on the left, and the monkey from the “fasting” group is on the right. There is a striking difference between these two macaques: the control animal has a hopeless look of a pessimist philosopher, its mouth is open, the corners of the mouth are mournfully dropped, the skin is shabby, and the tail is hanging lifelessly. At the same time, the food-restricted animal is looking at the camera with wide-open eyes, the mouth is tightly closed, the skin is in order, it has thick fur, and the tail is optimistically directed upwards.
Fig. 1. Rhesus macaques monkeys without food restriction (left) and with food restriction (right), age 28 years (after Colman et al. [30]).
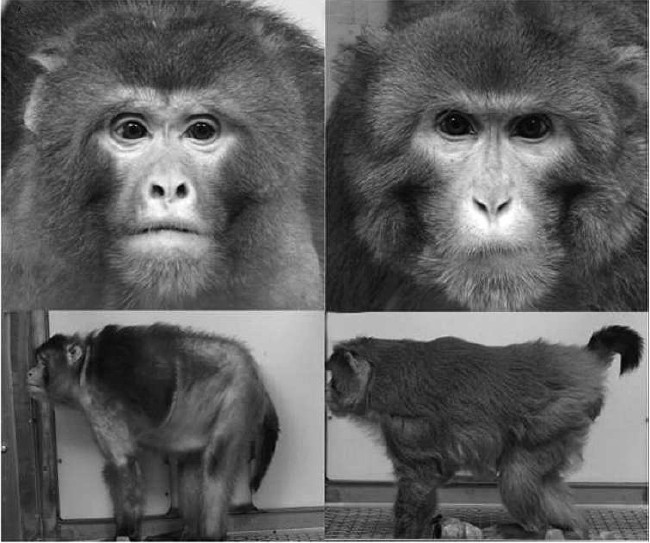
The experiment on monkeys is not completed yet, so we cannot draw conclusions on the effects of food restriction on the maximal lifespan of primates. However, such data are available for rodents. They show that in mice, rats, and hamsters the median lifespan increases much more than the maximal lifespan. Canceling or significant inhibition of the aging program provides the simplest explanation for the increase in the median lifespan. Other ontogenetic programs, primarily the growth of the organism, can also be slowed. This occurs with sufficiently strong and long starvation. However, more moderate food restriction can also prolong life without retardation of growth. With regard to maximal lifespan, its increase requires first of all abolition of all forms of cancer; otherwise, the organism with cancelled aging program will eventually get cancer. Both food restriction and our antioxidant SkQ have a protective effect in relation to certain cancer types, but unfortunately, these types seem to be in the minority [2, 32].
The effects of food restriction and SkQ proved to have much in common. In both cases, we observe rectangularization of mortality curves; early mortality is drastically reduced, and median lifespan increases much more than maximal lifespan. Their effect is mainly not due to the extension of life per se, but about the extension of healthy, youthful life. Both effects are manifested in the living beings that differ greatly in their systematic position (food restriction has geroprotective effect in yeast, worms, insects, and mammals; SkQ – in mycelial fungus, flowering plant, crustacean, insect, fish, and mammal). The effects of both food restriction and SkQ are of pronounced pleiotropic character, i.e. they cause responses in many physiological systems of the organism. Cardiovascular diseases, osteoporosis, disorders of the visual apparatus, some types of cancer recede; no signs of hair graying, balding, and senile depression are observed. Contradictory data were obtained on sarcopenia and immune response in case of food restriction. A number of authors note that this effect weakens both the muscular system and immunity [25, 33]. However, Weindruch et al. [30] indicate the absence of sarcopenia in monkeys, and MacCay, Robertson et al. [4-7] describe the resistance to lung infections in rats subjected to food restriction (compare to a sharp decrease in mortality from infectious diseases and inhibition of age-dependent thymus involution and follicular spleen compartments caused by SkQ1 [34, 35]). Food restriction and SkQ1 had opposite effects on wounds: starvation inhibited, and SkQ1, on the contrary, stimulated their healing [36]. Certain aspects of aging of rat visual apparatus could not be inhibited by food restriction [37], although SkQ1 proved to be very effective in this respect [10, 35]. Food restriction reduced body temperature and slowed the animals’ growth; these effects were not observed in case of SkQ1 [22]. All these data contradict such a trivial interpretation of our data as the assumption that SkQ1 reduces food consumption by animals due to reduced appetite. Direct measurements of food consumption by mice receiving SkQ1 have shown that SkQ1 has no effect on this parameter [38].
It is not surprising that food restriction caused deterioration of a number of vital parameters. When food is abundant, animals usually eat as much as they need to maintain vital functions. Therefore, continued food restriction leads to some disorders; the longer and more intense is starvation, the higher is probability of such disorders. We have already mentioned that geroprotective effect of food restriction does not require long and continuous starvation. For example, it is generally accepted that prolonged food restriction reduces the frequency of estrous cycles (sometimes leading to their complete disappearance) [39], but already in 1949 Carr et al. showed that temporal restriction followed by its removal, on the contrary, contributes to the preservation of estrous cycles until very old age (for reference see [2]). Note that a similar effect was also observed in the case of SkQ1 [34, 35, 38]. In general, there is no need for lifelong fasting if starvation is only a signal to prolong life by slowing the aging program [23].
There is another major factor that should be kept in mind when considering malnutrition as a geroprotector for humans. If this restriction is a danger signal, then the organism’s response should not be limited only to lifespan increase. Other responses are also highly probable, and some of them may be not nearly as attractive as extension of healthy life. For example, in a state of malnutrition a person becomes irritable and short-tempered, and a mouse, once in a “hamster wheel”, does not want to leave it, running from 6 to 8 km per night (this distance is always less than 1 km when the animal is normally fed), and it sometimes dies in the wheel from exhaustion [39]. This effect cannot be explained by a simple exhaustion caused by starvation. Rather, here is another response to the hunger signal: extreme concern and attempt to scan the largest possible area in search of food. If SkQ had a similar effect, we would observe an increase in food consumption by SkQ-treated animals [38]. It seems that SkQ administration is a “purer” way to slow the aging program, not burdened by undesirable side effects [3, 23].
In the coming months, we are planning clinical trials of the geroprotective effect of SkQ1 preparation on people taking it per os. With regard to food restriction, according to L. M. Redman and E. Ravussin [40], the only scientific result on its geroprotective effect on people was obtained back in 1957 by Argentinean researcher E. A. Vallejo [41]. Sixty people fasted every other day so that their average food consumption was reduced by 35%. Sixty people in the control group could eat without any restrictions. Apart from a brief report in 1957 [41], a detailed analysis of the data was published 18 years later by A. J. Stunkard and M. Rockstein [42], who reported a downward trend in mortality and 50% reduction of the average number of days spent in hospital in the people with restricted food consumption.
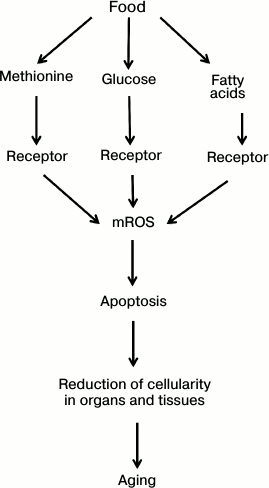
The concept presented in this section can be summed up in a scheme (see Fig. 2). It is postulated than an organism monitors the intake of three main food components (proteins, carbohydrates, and fats) using special receptors measuring blood concentration of methionine, glucose, and fatty acids, respectively. Receptors having bound these metabolites trigger a chain of events increasing the level of mitochondrial reactive oxygen species (mROS). In turn, mROS activate apoptosis, reducing organ and tissue cellularity, which weakens the organism’s functions, i.e. causes aging. Food restriction reduces the levels of methionine, glucose, and fatty acids and thus slows (or even cancels) the entire lethal cascade of senile phenoptosis shown in the figure.
Fig. 2. How eating ad libitum can shorten our life (explanation in text).
GEROPROTECTIVE EFFECT OF COLD
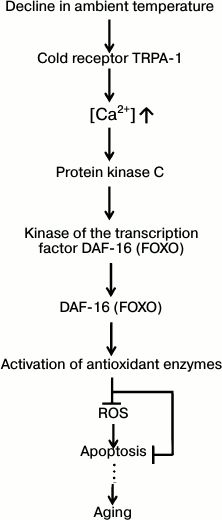
In the scientific literature, one can find a description of a number of cases when lowering the ambient temperature prolonged life (for review, see [43]). By analogy with our concept of the geroprotective effect of food restriction (Fig. 2), this phenomenon could be explained by an organism’s attempt to cancel the counterproductive aging program so as to support the organism’s functioning in deteriorating conditions (hypothermia). Recent experiments of R. Xiao et al. [44] on C. elegans (a poikilothermic animal, so lowering of the ambient temperature should slow all enzymatic processes) are especially demonstrative. It was shown that decline in ambient temperature from 25 to 15°C increases the median lifespan of this invertebrate from 9 to 30 days. Mutation-caused disabling of one of the worm’s genes, namely, the one encoding the protein of the calcium channel activated by cold (TRPA-1), significantly reduces the cold effect (in this case, at 15°C the worm lives for only 18 days instead of 30, while its lifespan at 25°C remains the same, 9 days). It is known that Ca2+ increase in the worm’s cells inactivates a protein-enzyme cascade acting in the following sequence: 1) protein kinase C; 2) kinase of the DAF-16 protein (one of the FOXO transcription factors), and 3) the actual DAF-16 protein. The latter regulates the activity of a large group of genes including intracellular proteins of antioxidant protection (activated by DAF-16) and apoptosis (inhibited), and the gene of extracellular superoxide dismutase-3 (activated). Successive knockout of genes of each protein in this cascade led to the disappearance of the part of the cold effect caused by the TRPA-1 protein. If, instead of knockout of the TRPA-1 protein gene, its activity was artificially increased, the cold effect also increased – worms now lived for 36 days at 15°C. Moreover, if the knocked out gene of the worm’s TRPA-1 protein was replaced by the gene of a homologous human protein, the regular cold effect on the nematode’s lifespan was restored. This observation indicates the universal character (from invertebrates to humans) of the newly discovered mechanism of inhibition of the aging program (Fig. 3).
Fig. 3. The way cold prolongs the lifespan of the nematode C. elegans (after Xiao et al. [44]; explanations in the text).
PAIN RECEPTOR STIMULATES THE AGING PROGRAM
On May 22, 2014, A. Dillin et al. from the University of California, Berkley published an important discovery in Cell that sheds light on one of the receptor mechanisms supporting the activity of the aging program [45]. They showed that knockout of the gene of one of the pain receptors, namely TRPV1 (Transient Receptor Potential cation channel subfamily V member 1), increases the lifespan of mice by 12-16%. TRPV1 is expressed in afferent sensory DRG (Dorsal Root Ganglion) neurons detecting very high temperature and pain stimuli in the dermal and epidermal skin layers, oral and nasal mucosa, joints, brain (where TRPV1 affects hippocampal synaptic plasticity), and several other tissues. These neurons transmit signals to the central nervous system via the spinal cord. DRG neurons are also found in the pancreas. Their demyelinated C-fibers expressing TRPV1 densely entwine β-cells of this gland. They secrete two types of neuropeptides: peptide P (stimulates inflammation) and peptide CGRP (Calcitonin Gene-Related Peptide, inhibits insulin production). The level of CGRP peptide was shown to increase dramatically with age [45, 46]. The mechanism of this effect remains unclear, but it is surely directly related to the development of type II diabetes due to the blockage of insulin secretion by β-cells [45]. According to M. S. Westerterp-Plantenga et al. [47], a diet rich in capsaicin (the substance obtained from hot pepper, an agonist of pain receptor TRPV1 and inhibitor of complex I similar in its action to rotenone) causes death of DRG neurons due to their overstimulation and reduces the number of cases of diabetes in people.
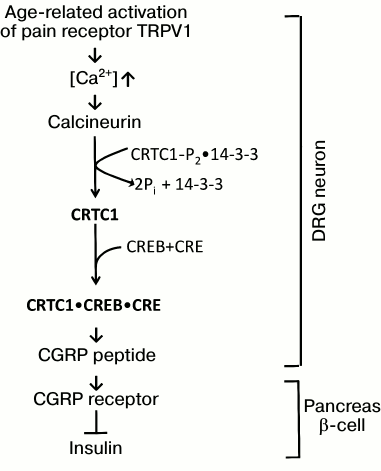
Figure 4 shows the chain of events from TRPV1 receptor activation to inhibition of insulin secretion discovered by Dillin et al. It includes: 1) increase in the level of intracellular Ca2+ penetrating through the neuron plasma membrane by a cationic channel of the TRPV1 receptor; 2) activation of the calcineurin protein; 3) calcineurin-mediated dephosphorylation of another protein, CRTC1 (CREB-Regulated Translational Coactivator 1), and, as a result, destruction of CRTC1 complex with 14-3-3 protein; 4) binding of released CRTC1 to transcriptional factors CREB and CRE; 5) CREB- and CRE-mediated activation of expression of the gene encoding CGPR peptide with subsequent CGRP synthesis and its secretion by nerve endings of DRG neurons into the intercellular space close to the pancreas β-cells; 6) CGRP binding by its receptor on the outer membrane of β-cells; 7) inhibition of insulin secretion by these cells.
Fig. 4. Activation of pain receptor in DRG neurons inhibits insulin production by pancreas β-cells (after Riera et al. [45]; explanations in the text).
Knockout of any of the proteins indicated in Fig. 4 halted the entire regulatory chain. In vivo administration for 6 weeks of peptide CGRP8-37 (a shortened version of CGRP, the CGRP receptor antagonist on β-cells) had the same effect. This compound prolonged the lifespan of mice and, similar to the TRPV1 gene knockout, inhibited the development of a number of senile signs in animals (in particular, reduction of O2 absorption in vivo, disappearance of the circadian rhythm in respiratory coefficient changes, decreased expression of insulin gene, weakening of spatial memory, impaired motor coordination) [45].
We believe that the scheme presented in Fig. 4 describes an essential part of the program of organism aging. The scheme does not include its beginning (what exactly causes age-related activation of TRPV1 receptor) and the end (what are the concrete consequences of the drastic decrease in insulin level). It is possible that age-related deterioration of the condition of joints causes pain, which triggers TRPV1 receptors present in joints. There is also another option – the “big biological clock” counting down our age gives a signal to stop the production of some juvenile hormone – an antagonist of TRPV1 receptor or, to the contrary, to stimulate the production of an aging hormone – agonist of this receptor. The hippocampus, which is controlled by TRPV1, could participate in these events. The hippocampus and its calcium homeostasis are known to be highly sensitive to aging [48-50].
Most likely, ROS serve as a necessary intermediate somewhere in the pathway leading from the hypothetical “clock” controlling ontogenesis and aging to the receptor. This can explain the geroprotective effect of cold receptor TRPA-1, opposing to the progeric effect of TRPV1. As we have already noted (see previous section), TRPA-1 activation stimulates a transcription factor of FOXO type, including transcription of intra- and extracellular antioxidant enzymes and antiapoptotic factors.
As for the events triggered by the lack of insulin, they are reversed by mitochondrial antioxidant SkQ1 (V. Skulachev, F. Severin, V. Popov, T. Popova et al., manuscript in preparation). This is consistent with the numerous data on the development of persistent oxidative stress in diabetes (see, for example, [51, 52]).
Generally, the more we learn about diabetes, the more it resembles the programs of aging, cancer, or septic shock [1-3], i.e. death orchestrated by the organism itself. In particular, overeating, characteristic for the elderly, should stimulate both aging (see above, Fig. 2) and diabetes. The program of septic shock with its sharp increase in fatty acid levels in blood [53, 54] could use the receptor of these acids to kill the organism via the same cascade as aging (Fig. 2), but in its significantly accelerated mode.
Concluding this section, we would like to emphasize that cancellation of aging in the naked mole rat is accompanied by the disappearance of CGRT peptide (and with it also diabetes as one of the causes of death) and desensitization of the organism to capsaicin [55]. Disappearance of the aging program combined with the development of antioxidant, anticancer, and antiapoptotic protection by high molecular weight hyaluronan and the greater accuracy in the work of ribosomes (for discussion, see [3]) could explain the extraordinary longevity of this mammal.
POSSIBLE PSYCHOLOGICAL ASPECTS AFFECTING HUMAN LIFESPAN
Consideration of aging as the program regulated by the organism in response to changing external conditions raises the question of the possibility for a particular person to actively fight their aging.
In 2013, one of the top scientific journals (PNAS) published an article by S. Cole et al. from the University of California Los Angeles entitled “A functional genomic perspective on human well-being” [56]. They measured gene expression in blood leukocytes of 84 Americans, dividing them into two groups based on the type of pleasure they get from life. The first type of pleasure is hedonistic; it is about simple satisfaction of one’s personal needs. The second type represents a deeper eudemonistic pleasure, which is related to the awareness of the importance of one’s existence not only for oneself, but also for others, self-improvement, and pursuit of some higher goals. Surprisingly, a statistically significant difference was observed between the two groups in gene expression of proteins involved in inflammatory reactions of the immune system: in hedonistic type, pleasure significantly increased this expression, and in eudemonistic type – to the contrary, reduced it.
C. Franceschi considers chronic inflammation as one of the main causes of aging (“inflamm-aging”) [57]. This process is known to play an important role in the development of cancer [57]. That is why it would be logical to assume that adherents of hedonistic pleasures will live shorter lives than those choosing eudemonistic pleasures.
This logic could explain a striking observation made recently by V. A. Anisimov and G. M. Zharinov. They compared the average lifespans of musicians playing classical music with those of rock musicians. This was shown to be 70 years for male violinists, and only 45 years for rock musicians. The difference was even bigger for women – 78 against 36.5 years [58, 59]. These figures might be explained by different way of life of classical and rock musicians, but such an explanation is not enough to understand why the average lifespan of female harpists is 81 years and that of female pianists is 74.5 years. (On therapeutic and biochemical effects of classical music, see [60-63]).
The most recent studies have shown that phenoptotic aging program is controlled by receptor systems that monitor the key parameters of the external and internal environment of the organism. Under unfavorable conditions, when it comes to the survival of the population, the organism can slow down this counterproductive program, reducing evolvability but increasing the chances of individual survival. We can assume that in the case of humans, deceleration of the aging program will be observed in individuals acting for the benefit of communities.
According to Penninx et al. [64], among the many psychological factors, there are two most clearly correlated with longevity: awareness of one’s usefulness to others and maintaining high level of excellence in old age. As for the individuals harmful for the community, they can be eliminated by acute phenoptosis. In the 1920s, cases were recorded when following the shaman’s order, a community of Australian aborigines started singing funeral chants for a still living person, inducing disorders of salt metabolism that killed the victim in just a few days [65]. On the other hand, the geroprotective effect of classical music could be explained by its opposite (i.e. inhibiting) effect on phenoptotic programs. In this context, an important task of gerontology is to elucidate the entire list of factors that stimulate or inhibit the aging program. Understanding of these factors might help one to master his destiny.
In any case, the very fact of adjustability of age-related pathologies by the organism’s control systems is a very strong argument in favor of the concept of aging as a program. It greatly increases the probability of success in deceleration of aging by pharmaceutical compounds inhibiting such a program, for example, mitochondria-addressed antioxidants.
The study is financially supported by the Russian Science Foundation (grant 14-24-00107).
REFERENCES
1.Skulachev, V. P. (2003) Aging and the programmed
death phenomena, in Topics in Current Genetics, Model Systems
in Aging (Nystrom, T., and Osiewacz, H. D., eds.) Springer-Verlag,
Berlin-Heidelberg, pp. 192-237.
2.Skulachev, V. P., Skulachev, M. V., and Feniuk, B.
A. (2013) Life without Aging [in Russian], EKSMO, Moscow.
3.Skulachev, M. V., and Skulachev, V. P. (2014) New
data on programmed aging – slow phenoptosis, Biochemistry
(Moscow), 79, 977-993.
4.MacCay, C. M., and Crowell, M. F. (1934) Prolonging
the life span, Sci. Mon., 39, 405-414.
5.Robertson, T. B., Marston, R., and Walters, J. W.
(1934) Influence of intermittent starvation and of intermittent
starvation plus nucleic acid on growth and longevity in white mice,
Aust. J. Exp. Biol. Med. Sci., 12, 33.
6.MacCay, C. M., Crowell, M. F., and Maynard, L. A.
(1935) The effect of retarded growth upon the length of life span and
upon the ultimate body size, J. Nutr., 10, 63-79.
7.MacCay, C. M., Maynard, L. A., and Barnes, L. L.
(1943) Growth, aging, chronic disease and lifespan in rats, Arch.
Biochem., 2, 469.
8.Harman, D. (1956) Aging: a theory based on free
radical and radiation chemistry, J. Gerontol., 11,
298-300.
9.Will, L. C., and McCay, C. M. (1943) Ageing, basal
metabolism and retarded growth, Arch. Biochem., 2,
481.
10.Mair, W., Goymer, P., Pletcher, S. D., and
Partridge, L. (2003) Demography of dietary restriction and death in
Drosophila, Science, 301, 1731-1733.
11.Anisimov, V. N., Bakeeva, L. E., Egormin, P. A.,
Filenko, O. F., Isakova, E. F., Manskikh, V. N., Mikhelson, V. M.,
Panteleeva, A. A., Pasyukova, E. G., Pilipenko, D. I., Piskunova, T.
S., Popovich, I. G., Roshchina, N. V., Rybina, O. Yu., Saprunova, V.
V., Samoylova, T. A., Semenchenko, A. V., Skulachev, M. V., Spivak, I.
M., Tsybul’ko, E. A., Tyndyk, M. L., Vyssokikh, M. Yu., Yurova,
M. N., Zabezhinsky, M. A., and Skulachev, V. P. (2008)
Mitochondria-targeted plastoquinone derivatives as tools to interrupt
execution of the aging program. 5. SkQ1 prolongs lifespan and prevents
development of traits of senescence, Biochemistry (Moscow),
73, 1329-1342.
12.Libert, S., and Pletcher, S. D. (2007) Modulation
of longevity by environmental sensing, Cell, 131,
1231-1234.
13.Libert, S., Zwiener, J., Chu, X., Vanvoorhies,
W., Roman, G., and Pletcher, S. D. (2007) Regulation of Drosophila
life span by olfaction and food-derived odors, Science,
315, 1133-1137.
14.Stuchlikova, E., Juricova-Horakova, M., and Deyl,
Z. (1975) New aspects of the dietary effect of life prolongation in
rodents. What is the role of obesity in aging? Exp. Gerontol.,
10, 141-144.
15.Richie, J. P., Jr., Leutzinger, Y.,
Parthasarathy, S., Malloy, V., Orentreich, N., and Zimmerman, J. A.
(1994) Methionine restriction increases blood glutathione and longevity
in F344 rats, FASEB J., 8, 1302-1307.
16.Miller, R. A., Buehner, G., Chang, Y., Harper, J.
M., Sigler, R., and Smith-Wheelock, M. (2005) Methionine-deficient diet
extends mouse lifespan, slows immune and lens aging, alters glucose,
T4, IGF-I and insulin levels, and increases hepatocyte MIF levels and
stress resistance, Aging Cell, 4, 119-125.
17.Sanz, A., Caro, P., Ayala, V., Portero-Otin, M.,
Pamplona, R., and Barja, G. (2006) Methionine restriction decreases
mitochondrial oxygen radical generation and leak as well as oxidative
damage to mitochondrial DNA and proteins, FASEB J., 20,
1064-1073.
18.Caro, P., Gomez, J., Sanchez, I., Garcia, R.,
Lopez-Torres, M., Naudi, A., Portero-Otin, M., Pamplona, R., and Barja,
G. (2009) Effect of 40% restriction of dietary amino acids (except
methionine) on mitochondrial oxidative stress and biogenesis, AIF and
SIRT1 in rat liver, Biogerontology, 10, 579-592.
19.Sanchez-Roman, I., Gomez, A., Perez, I., Sanchez,
C., Suarez, H., Naudi, A., Jove, M., Lopez-Torres, M., Pamplona, R.,
and Barja, G. (2012) Effects of aging and methionine restriction
applied at old age on ROS generation and oxidative damage in rat liver
mitochondria, Biogerontology, 13, 399-411.
20.Sanchez-Roman, I., Gomez, A., Gomez, J., Suarez,
H., Sanchez, C., Naudi, A., Ayala, V., Portero-Otin, M., Lopez-Torres,
M., Pamplona, R., and Barja, G. (2011) Forty percent methionine
restriction lowers DNA methylation, complex I ROS generation, and
oxidative damage to mtDNA and mitochondrial proteins in rat heart,
J. Bioenerg. Biomembr., 43, 699-708.
21.Edman, U., Garcia, A. M., Busuttil, R. A.,
Sorensen, D., Lundell, M., Kapahi, P., and Vijg, J. (2009) Lifespan
extension by dietary restriction is not linked to protection against
somatic DNA damage in Drosophila melanogaster, Aging
Cell, 8, 331-338.
22.Skulachev, V. P. (2011) SkQ1 treatment and food
restriction – two ways to retard an aging program of organisms,
Aging (Albany), 3, 1045-1050.
23.Skulachev, V. P. (2012) What is
“phenoptosis” and how to fight it? Biochemistry
(Moscow), 77, 689-706.
24.Remolina, S. C., and Hughes, K. A. (2008)
Evolution and mechanisms of long life and high fertility in queen honey
bees, Age (Dordr.), 30, 177-185.
25.Sun, D., Muthukumar, A. R., Lawrence, R. A., and
Fernandes, G. (2001) Effects of calorie restriction on polymicrobial
peritonitis induced by cecum ligation and puncture in young C57BL/6
mice, Clin. Diagn. Lab. Immunol., 8, 1003-1011.
26.Garcia, A. M., Busuttil, R. A., Calder, R. B.,
Dolle, M. E., Diaz, V., McMahan, C. A., Bartke, A., Nelson, J.,
Reddick, R., and Vijg, J. (2008) Effect of Ames dwarfism and caloric
restriction on spontaneous DNA mutation frequency in different mouse
tissues, Mech. Ageing Dev., 129, 528-533.
27.Hamden, K., Carreau, S., Ayadi, F., Masmoudi, H.,
and El Feki, A. (2009) Inhibitory effect of estrogens, phytoestrogens,
and caloric restriction on oxidative stress and hepato-toxicity in aged
rats, Biomed. Environ. Sci., 22, 381-387.
28.Park, S. K., Kim, K., Page, G. P., Allison, D.
B., Weindruch, R., and Prolla, T. A. (2009) Gene expression profiling
of aging in multiple mouse strains: identification of aging biomarkers
and impact of dietary antioxidants, Aging Cell, 8,
484-495.
29.Eng, P. M., Rimm, E. B., Fitzmaurice, G., and
Kawachi, I. (2002) Social ties and change in social ties in relation to
subsequent total and cause-specific mortality and coronary heart
disease incidence in men, Amer. J. Epidemiol., 155,
700-709.
30.Colman, R. J., Anderson, R. M., Johnson, S. C.,
Kastman, E. K., Kosmatka, K. J., Beasley, T. M., Allison, D. B.,
Cruzen, C., Simmons, H. A., Kemnitz, J. W., and Weindruch, R. (2009)
Caloric restriction delays disease onset and mortality in rhesus
monkeys, Science, 325, 201-204.
31.Colman, R. J., and Anderson, R. M. (2011)
Nonhuman primate calorie restriction, Antioxid. Redox Signal.,
14, 229-239.
32.Severina, I. I., Severin, F. F., Korshunova, G.
A., Sumbatyan, N. V., Ilyasova, T. M., Simonyan, R. A., Rogov, A. G.,
Trendeleva, T. A., Zvyagilskaya, R. A., Dugina, V. B., Domnina, L. V.,
Fetisova, E. K., Lyamzaev, K. G., Vyssokikh, M. Y., Chernyak, B. V.,
Skulachev, M. V., Skulachev, V. P., and Sadovnichii, V. A. (2013) In
search of novel highly active mitochondria-targeted antioxidants:
thymoquinone and its cationic derivatives, FEBS Lett.,
587, 2018-2024.
33.Gardner, E. M. (2005) Caloric restriction
decreases survival of aged mice in response to primary influenza
infection, J. Gerontol. A. Biol. Sci. Med. Sci., 60,
688-694.
34.Obukhova, L. A., Skulachev, V. P., and Kolosova,
N. G. (2009) Mitochondria-targeted antioxidant SkQ1 inhibits
age-dependent involution of the thymus in normal and senescence-prone
rats, Aging, 1, 389-401.
35.Skulachev, V. P., Anisimov, V. N., Antonenko, Y.
N., Bakeeva, L. E., Chernyak, B. V., Erichev, V. P., Filenko, O. F.,
Kalinina, N. I., Kapelko, V. I., Kolosova, N. G., Kopnin, B. P.,
Korshunova, G. A., Lichinitser, M. R., Obukhova, L. A., Pasyukova, E.
G., Pisarenko, O. I., Roginsky, V. A., Ruuge, E. K., Senin, I. I.,
Severina, I. I., Skulachev, M. V., Spivak, I. M., Tashlitsky, V. N.,
Tkachuk, V. A., Vyssokikh, M. Y., Yaguzhinsky, L. S., and Zorov, D. B.
(2009) An attempt to prevent senescence: a mitochondrial approach,
Biochim. Biophys. Acta, 1787, 437-461.
36.Demianenko, I. A., Vasilieva, T. V., Domnina, L.
V., Dugina, V. B., Egorov, M. V., Ivanova, O. Y., Ilinskaya, O. P.,
Pletjushkina, O. Y., Popova, E. N., Sakharov, I. Y., Fedorov, A. V.,
and Chernyak, B. V. (2010) Novel mitochondria-targeted antioxidants,
“Skulachev-ion” derivatives, accelerate dermal wound
healing in animals, Biochemistry (Moscow), 75,
274-280.
37.Obin, M., Halbleib, M., Lipman, R., Carroll, K.,
Taylor, A., and Bronson, R. (2000) Calorie restriction increases
light-dependent photoreceptor cell loss in the neural retina of Fischer
344 rats, Neurobiol. Aging, 21, 639-645.
38.Anisimov, V. N., Egorov, M. V., Krasilshchikova,
M. S., Lyamzaev, K. G., Manskikh, V. N., Moshkin, M. P., Novikov, E.
A., Popovich, I. G., Rogovin, K. A., Shabalina, I. G., Shekarova, O.
N., Skulachev, M. V., Titova, T. V., Vygodin, V. A., Vyssokikh, M. Y.,
Yurova, M. N., Zabezhinsky, M. A., and Skulachev, V. P. (2011) Effects
of the mitochondria-targeted antioxidant SkQ1 on lifespan of rodents,
Aging (Albany NY), 3, 1110-1119.
39.Hopkin, K. (2003) Dietary drawbacks, Sci.
Aging. Knowl. Environ., 2003, No. 8, p. NS4.
40.Redman, L. M., and Ravussin, E. (2011) Caloric
restriction in humans: impact on physiological, psychological, and
behavioral outcomes, Antiox. Redox Signal., 14,
275-287.
41.Vallejo, E. A. (1957) Hunger diet on alternate
days in the nutrition of the aged, Prensa Med. Argent.,
44, 119-120.
42.Stunkard, A. J., and Rockstein, M. (1976)
Nutrition, longevity and obesity, in Nutrition, Aging and
Obesity (Rockstein, M., and Stunkard, A. J., eds.) Academic Press,
N.Y., pp. 253-284.
43.Mitteldorf, J., and Sagan, D. (2014) Suicide
Genes, MacMillan Press, in press.
44.Xiao, R., Zhang, B., Dong, Y. M., Gong, J. K.,
Xu, T., Liu, J. F., and Xu, X. Z. S. (2013) A genetic program promotes
C. elegans longevity at cold temperatures via a thermosensitive
TRP channel, Cell, 152, 806-817.
45.Riera, C. E., Huising, M. O., Follett, P.,
Leblanc, M., Halloran, J., Van Andel, R., de Magalhaes Filho, C. D.,
Merkwirth, C., and Dillin, A. (2014) TRPV1 pain receptors regulate
longevity and metabolism by neuropeptide signaling, Cell,
157, 1023-1036.
46.Melnyk, A., and Himms-Hagen, J. (1995) Resistance
to aging-associated obesity in capsaicin-desensitized rats one year
after treatment, Obes. Res., 3, 337-344.
47.Westerterp-Plantenga, M. S., Smeets, A., and
Lejeune, M. P. (2005) Sensory and gastrointestinal satiety effects of
capsaicin on food intake, Int. J. Obes. (Lond.), 29,
682-688.
48.Moreno, H., Burghardt, N. S., Vela-Duarte, D.,
Masciotti, J., Hua, F., Fenton, A. A., Schwaller, B., and Small, S. A.
(2012) The absence of the calcium-buffering protein calbindin is
associated with faster age-related decline in hippocampal metabolism,
Hippocampus, 22, 1107-1120.
49.West, M. J. (1993) Regionally specific loss of
neurons in the aging human hippocampus, Neurobiol. Aging,
14, 287-293.
50.West, M. J., Coleman, P. D., Flood, D. G., and
Troncoso, J. C. (1994) Differences in the pattern of hippocampal
neuronal loss in normal ageing and Alzheimer’s disease,
Lancet, 344, 769-772.
51.Rains, J. L., and Jain, S. K. (2011) Oxidative
stress, insulin signaling, and diabetes, Free Radic. Biol. Med.,
50, 567-575.
52.Brownlee, M. (2001) Biochemistry and molecular
cell biology of diabetic complications, Nature, 414,
813-820.
53.Maitra, U., Chang, S., Singh, N., and Li, L.
(2009) Molecular mechanism underlying the suppression of lipid
oxidation during endotoxemia, Mol. Immunol., 47,
420-425.
54.Schaffler, A., and Scholmerich, J. (2010) Innate
immunity and adipose tissue biology, Trends Immunol., 31,
228-235.
55.Park, T. J., Comer, C., Carol, A., Lu, Y., Hong,
H. S., and Rice, F. L. (2003) Somatosensory organization and behavior
in naked mole-rats. II. Peripheral structures, innervation, and
selective lack of neuropeptides associated with thermoregulation and
pain, J. Comp. Neurol., 465, 104-120.
56.Fredrickson, B. L., Grewen, K. M., Coffey, K. A.,
Algoe, S. B., Firestine, A. M., Arevalo, J. M., Ma, J., and Cole, S. W.
(2013) A functional genomic perspective on human well-being, Proc.
Natl. Acad. Sci. USA, 110, 13684-13689.
57.Franceschi, C., Bonafe, M., Valensin, S.,
Olivieri, F., De Luca, M., Ottaviani, E., and De Benedictis, G. (2000)
Inflamm-aging. An evolutionary perspective on immunosenescence, Ann.
New York Acad. Sci., 908, 244-254.
58.Zharinov, G., and Anisimov, V. (2014) Classical
music instead of an anti-aging pill? Skripichnyi Kluch,
1, 33-36.
59.Anisimov, V. N., and Zharinov, G. M. (2013) Muses
and longevity, Geront. Geriatr. Res., 2, 1000e1123.
60.Sharma, D. C. (2013) Impact analysis of sounds of
nature (music therapy) on the hemato-biochemistry of Homo
sapiens and Rattus norvegicus, in 2nd Int. Conf. Adv.
Biol. Pharmac. Sci., Hong Kong, pp. 6-8.
61.Gangrade, A. (2012) The effect of music on the
production of neurotransmitters, hormones, cytokines, and peptide: a
review, Music and Medicine, 4, 40-43.
62.Da Cruz, J. N., De Lima, D. D., Dal Magro, D. D.,
and Da Cruz, J. G. P. (2011) The power of classic music to reduce
anxiety in rats treated with Simvastatin, Basic Clin. Neurosci.,
2, 5-11.
63.Uchiyama, M., Jin, X. Y., Zhang, Q., Hirai, T.,
Amano, A., Bashuda, H., and Niimi, M. (2012) Auditory stimulation of
opera music induced prolongation of murine cardiac allograft survival
and maintained generation of regulatory CD4(+)CD25(+) cells, J.
Cardiothorac. Surg., 7, 1-8.
64.Penninx, B. W., van Tilburg, T., Kriegsman, D.
M., Deeg, D. J., Boeke, A. J., and van Eijk, J. T. (1997) Effects of
social support and personal coping resources on mortality in older age:
the longitudinal aging study Amsterdam, Am. J. Epidemiol.,
146, 510-519.
65.Eastwell, H. D. (1987) Voodoo death in Australian
aborigines, Psychiatr. Med., 5, 71-73.