Preventive and Therapeutic Effects of SkQ1-Containing Visomitin Eye Drops against Light-Induced Retinal Degeneration
Yu. P. Novikova1, O. S. Gancharova2*, O. V. Eichler2, P. P. Philippov2, and E. N. Grigoryan1*
1Koltzov Institute of Developmental Biology, 119334 Moscow, Russia; fax: (499) 135-8012; E-mail: leonore@mail.ru2Lomonosov Moscow State University, Belozersky Institute of Physico-Chemical Biology, 119991 Moscow, Russia; fax: (495) 939-0978; E-mail: fxb@genebee.msu.su; olgancharova@gmail.com
* To whom correspondence should be addressed.
Received May 16, 2014
The human retina is constantly affected by light of varying intensity, this being especially true for photoreceptor cells and retinal pigment epithelium. Traditionally, photoinduced damages of the retina are induced by visible light of high intensity in albino rats using the LIRD (light-induced retinal degeneration) model. This model allows study of pathological processes in the retina and the search for retinoprotectors preventing retinal photodamage. In addition, the etiology and mechanisms of retina damage in the LIRD model have much in common with the mechanisms of the development of age-related retinal disorders, in particular, with age-related macular degeneration (AMD). We have studied preventive and therapeutic effects of Visomitin eye drops (based on the mitochondria-targeted antioxidant SkQ1) on albino rat retinas damaged by bright light. In the first series of experiments, rats receiving Visomitin for two weeks prior to illumination demonstrated significantly less expressed atrophic and degenerative changes in the retina compared to animals receiving similar drops with no SkQ1. In the second series, the illuminated rats were treated for two weeks with Visomitin or similar drops without SkQ1. The damaged retinas of the experimental animals were repaired much more effectively than those of the control animals. Therefore, we conclude that Visomitin SkQ1-containing eye drops have pronounced preventive and therapeutic effects on the photodamaged retina and might be recommended as a photoprotector and a pharmaceutical preparation for the treatment of AMD in combination with conventional medicines.
KEY WORDS: retina, photoreceptors, photodamage, SkQ1, VisomitinDOI: 10.1134/S0006297914100113
Abbreviations: AMD, age-related macular degeneration; GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; LIRD, light-induced retinal degeneration; ONL, outer nuclear layer; OPL, outer plexiform layer; PL, photoreceptor layer; RPE, retinal pigment epithelium; ROS, reactive oxygen species.
Reactive oxygen species (ROS), which are mainly generated by
mitochondrial electron transport [1-3], can cause considerable damage to various cell
types. This is especially true for the retina, which is exposed to
photon flux during the entire life of an organism, and in addition its
cells are characterized by high rate of oxygen metabolism and high
content of polyunsaturated fatty acids [4, 5]. Thus, conditions are created for regular
production of a large amount of ROS that damage retinal structures,
especially photoreceptor cells – the rods and cones [6-11]. It has been demonstrated
on many in vitro and in vivo models that the penetrating
cation 10-(6′-plastoquinonyl)decyltriphenylphosphonium (SkQ1) has
high antioxidant activity due to its ability to remove ROS from the
mitochondrial matrix [12-14].
According to our earlier data [15, 16], the antioxidant SkQ1 has protective effect on the neural retina, retinal pigment epithelium (RPE), and choroid cultivated in preparations of the posterior segment of albino rat eyes, significantly reducing cell death in these structures and preventing macrophage transformation of RPE cells. In the present study we examined the ability of Visomitin SkQ1-containing eye drops to provide prophylactic and therapeutic effects on albino rat retina damaged by exposure to high intensity light according to the LIRD (light-induced retinal degeneration) model [17, 18].
MATERIALS AND METHODS
Visomitin eye drops were obtained from the Institute of Mitoengineering, Lomonosov Moscow State University. The composition of the eye drops (per 100 ml): 0.0155 mg SkQ1, 10 mg benzalkonium chloride, 200 mg oxypropyl methylcellulose, 900 mg sodium chloride, 810 mg sodium dihydrophosphate, 1.1635 g sodium hydrogen phosphate dodecahydrate, and water to total volume 100 ml.
Animals. Healthy two-month-old female albino Wistar rats (n = 36) were used for the experiments, six of them serving as controls. Prior to the experiment, the animals were kept under normal vivarium conditions (12 h light/12 h dark); we conducted regular external inspection of their eyes. Upon completion of the experiment, the rats were decapitated under chloral hydrate (Sigma, USA) anesthesia (500 mg/kg intraperitoneally). All the procedures were consistent with the protocols approved by the Russian Academy of Sciences Commission on Bioethics and performed in compliance with the ethical rules of the European Directive FELASA-2010.
Damaging of albino rat retina using the LIRD model. Before illumination, the rats were kept in the dark for 14 h for dark adaptation of their eyes, and then they were exposed to bright light for 14 h. The high-intensity visible light source was an NC-DE 70W/DW RX7s metal halide lamp (NARVA, Germany) with the following specifications: power 70 W, luminous flux 5000-5500 lumens, color temperature 4000 K. The lamp was placed 2 m from the animal cages, which corresponded to intensity of retina illumination of 2500-3000 lx.
Preventive effect of Visomitin SkQ1-containing eye drops on albino rat retina. Rats (n = 18) of the experimental (n = 9) and control (n = 9) groups received, respectively, SkQ1-containing eye drops or similar drops without SkQ1 (placebo) for 15 days; in both cases, one drop per day was placed in the left and right eyes of each animal. Upon completion of the course, the animals were illuminated with the light of high intensity, and then were kept in the dark for 3 h. Immediately thereafter, six rats from the experimental and six rats from the control groups were euthanized. The remaining animals (three from each group) were kept for 15 days under the usual conditions (without illumination or administration of the eye drops) and then were euthanized.
Therapeutic effect of Visomitin SkQ1-containing eye drops on albino rat retina. The rats (n = 12) were illuminated with the high-intensity light and then kept for 15 days under the usual light conditions. The experimental group (n = 6) of illuminated animals received SkQ1-containing eye drops, and the control group (n = 6) received drops of the same composition without SkQ1 (placebo); in both cases the animals received one drop per day in the left and right eyes. Upon completion of the 15-days course of administration of the eye drops, the animals were euthanized.
Histological processing. Eyeballs isolated immediately after decapitation of the animals were placed in a Bouin’s fixative solution, and after two days the posterior segment containing retina, choroid, and sclera was isolated. Fixed preparations of the eye posterior segment were then washed in 70% alcohol and were subjected to routine histological processing including dehydration in alcohols of increasing concentration and embedding the properly oriented samples in paraffin. Sagittal serial sections (500 from each eye, 7-µm thick each) were obtained from the paraffin blocks using a microtome (Reichert, Austria). The sections fixed on slides (Menzel-Glaser, Germany) were deparaffinized, rehydrated, stained with Carazzi’s hematoxylin and eosin following the standard protocol, and mounted in Canada balsam.
Morphometric analysis. Serial histological preparations were studied using Vanox AH-3 (Olympus, Japan), Jenaval (Carl Zeiss, Germany), and AxioScope A.1 (Carl Zeiss) microscopes. Microphotographs were obtained using an AxioCam MRc5 high-resolution digital camera (Carl Zeiss). Viewing and processing of the microphotographs, addition of scales, and morphometric measurements were performed using the AxioVision 8.0 (Carl Zeiss) and Adobe Photoshop CS6 Extended (Adobe Systems, USA) softwares.
The width of the outer nuclear retinal layer (ONL) was examined 3 h after photodamaging. To estimate the change in this parameter caused by the intense light, two retinal preparations were randomly chosen from each group of rats euthanized 3 h after photodamaging. Five-hundred measurements of ONL width were carried out in 60 serial sections from each such preparation, in eight different areas from the dorsal to the ventral sides of the eye (two dorso-peripheral areas, two dorso-central areas, two ventro-central areas, and two ventro-peripheral areas).
ONL atrophic area size was evaluated 15 days after the high-intensity illumination. For this, two retinal preparations with the most severe damage were selected from each group of rats euthanized 15 days after illumination. ONL atrophic area size was determined as the product of the number of consecutive sections with detected ONL atrophy, section width (7 μm) and the length of the atrophy zone (in μm) in each section.
The length of the inner segments of photoreceptor cells was determined in the area of the highest light-sensitivity on 60 sections randomly selected from each group. The data were processed to determine the statistically significant differences of sample mean values using the Mann–Whitney test at p = 0.05. Results were calculated and plotted using the MS Office Excel and ORIGIN 8.1 programs.
RESULTS
Retinal damage caused by bright light. The structure of the retina of intact albino rats (n = 6) was typical for healthy tissue [19]. Figure 1 shows that the neural retina (following the direction from its scleral to vitreal parts) consists of three nuclear layers containing neuronal cell bodies that are interleaved with two plexiform layers consisting of neuronal processes. The layers interacting with RPE are located closer to the sclera; they include the outer segments and bodies of photoreceptor cells that together form the outer retina. The outer plexiform layer, inner nuclear layer (INL) formed of interneuron bodies, and ganglion cell layer are located more vitreally. The inner plexiform layer is located between INL and ganglion cell layer, which together form the inner retina.
Fig. 1. Examples of early photoinduced retinal damage in albino rats 3 h after exposure to bright light. a) Intact retina; b) ONL disorganization and thinning; c) ONL cells migration in the vitreal direction (white arrows); d) migration of an activated RPE cell in the vitreal direction (black arrow). Retinal cross sections; staining with hematoxylin and eosin; ×1000 magnification; scale bar 20 μm.
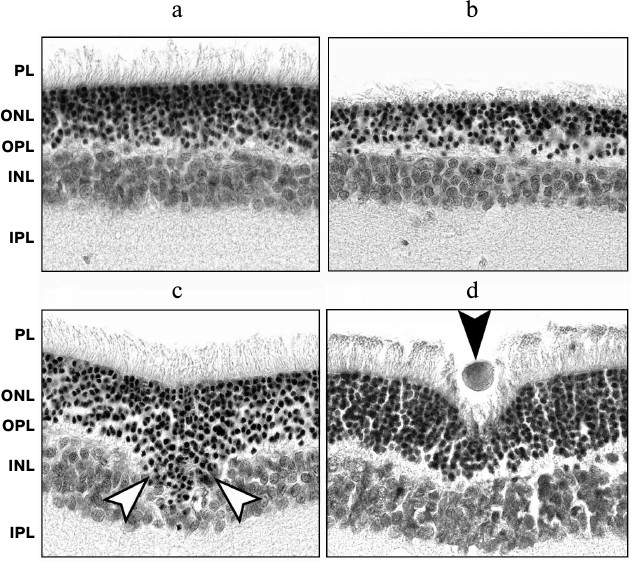
Damages typical for the LIRD models [17, 18, 20-26] were found in the retina of albino rats exposed to illumination with bright light (n = 9). Usually, morphological manifestations of early photoinduced damages typical for this model are found in the outer retina 2-4 h after illumination. They include destruction of the photoreceptor layer, the initial stage of ONL thinning, pyknosis of photoreceptor nuclei, as well as vacuolization, migration, and phagocytic activity of RPE cells [17, 27, 28]. In our case, shortening and disorganization of outer and inner segments of photoreceptor cells and pyknosis of their nuclei were detected 3 h after photodamaging. Such changes in photoreceptor chromatin indicating the beginning of apoptosis caused the decrease in ONL thickness, thinning of the outer plexiform retinal layer formed by photoreceptor axons, and shifting of the bodies of photoreceptor cells in the scleral direction. In addition, some RPE cells migrated in the vitreal direction (Fig. 1) and phagocytized the damaged segments of photoreceptor cells (not shown). In some cases the neural retina was peeled off the RPE and choroid, demonstrating bucklings (not shown) due to accumulation of edema fluid and cellular debris underneath.
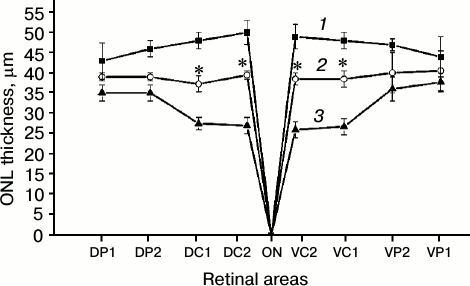
All the observed damages of the outer retina were of multifocal nature, which indicates unequal photosensitivity of its different areas. ONL thinning detected 3 h after photodamaging showed that dorso-central retinal area was most susceptible (Fig. 2).
Fig. 2. Comparison of ONL thickness before illumination (1) and 3 h after illumination in different eye areas of rats preventively treated with Visomitin eye drops (2) or similar eye drops without SkQ1 (3). ONL thickness was measured in the following retinal areas: ventro-peripheral (VP1, VP2), dorso-peripheral (DP1, DP2), ventro-central (VC1, VC2), and dorso-central (DC1, DC2). ONL is absent in the area of the optic nerve (ON) exit and its thickness is zero. Each curve represents data obtained for two retinal preparations.
Relatively late photoinduced damages typical for the LIRD model are manifested in albino rat retinas several days to several weeks after exposure to bright light. These changes are present in outer retinal layers in the form of subtotal ONL atrophy due to apoptotic death of photoreceptors and in inner retinal layers. In the latter case, it is manifested in the formation of microneuromas and channels and shifting of INL neurons in the scleral direction, resulting in significant general restructuring of the retina [17, 18, 20-24, 26]. In our experiments, 15 days after the illumination all the animals developed pronounced atrophic changes in ONL and retinal plexiform layers as well as accompanying migration of ONL and INL cells in the vitreal and scleral directions, respectively (Fig. 3). Atrophic loci were also observed in the RPE layer (not shown).
Fig. 3. Examples of late photoinduced retinal damages in albino rats 15 days after illumination with bright light. a, d) Intact retina; b) atrophy of outer and inner segments (PL) and bodies (ONL) of photoreceptor cells, shift of INL interneurons in the vitreal direction (arrow); c) reactive gliosis (hypertrophic processes of the Müller glia are indicated by arrows); e) retinal microneuroma (black arrow); f) severe disorders of retinal cytoarchitectonics; g) channel in the inner retina (white arrow) and retinal microneuroma (black arrow). Retinal cross sections; staining with hematoxylin and eosin; magnification ×1000 (a-c) and ×400 (d-g); scale bar, 20 μm.
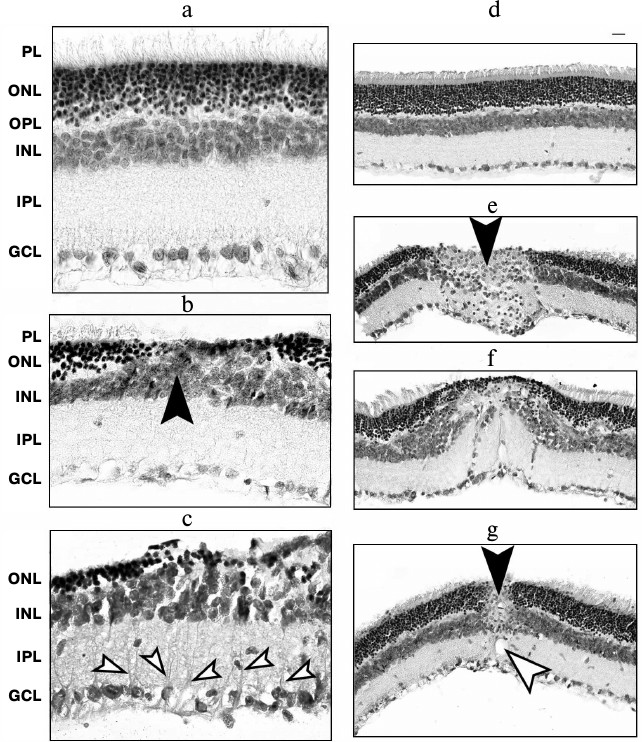
In addition, compensatory processes involving primarily the Müller glia of the neural retina developed in the albino rat retina during 15 days after its illumination with bright light. For example, in retinal areas adjacent to the areas of ONL atrophy, hypertrophy of glial cells processes could be observed that was manifested in increase in their thickness and number. Cysts and channels for draining the edema fluid were developing mainly in the INL. Microneuromas were detected in the INL – the clusters of neuronal-phenotype cells and randomly arranged bundles of their processes (Fig. 3). Complete destruction of retinal cytoarchitectonics up to indistinguishability of its layers could be observed in the most severely affected retinal areas.
Photodamage-associated changes of the ganglion cell layer (GCL) were less expressed, but death of cells of this layer was observed in about one third of the retinal samples obtained from the illuminated animals.
All the detected late retinal changes were of multifocal character; damaged areas in the inner retina usually accompanied the atrophy and other damages in the outer retina.
Preventive effect of Visomitin SkQ1-containing eye drops on retina of albino rats subjected to illumination with bright light. In this series of the experiments, albino rats received Visomitin SkQ1-containing drops (experimental group) or similar drops without SkQ1 (control group) for two weeks prior to the illumination. The state of retina in the experimental and control groups was compared after 3 h and 15 days to determine whether Visomitin had a preventive effect on the retina of illuminated animals. The severity of early and late pathological processes in the retina, typical for LIRD models, served as the criteria for this comparison.
Death of photoreceptors whose bodies are located in the ONL, resulting in the reduction of this layer thickness, is one of the earliest events of light-induced retinal damage. Comparison of ONL thickness in different areas of the retinal preparations (n = 2) obtained from animals of experimental and control groups, as well as from the intact rats, showed that ONL thickness was significantly greater in the experimental group (animals receiving Visomitin) than in the control group 3 h after illumination (Fig. 2), the effect of Visomitin being most pronounced in the central area of the retina. The degree of preservation of the outer and inner segments of photoreceptor cells, evaluated based on their size 3 h after illumination, was shown to be greater in rats from the experimental group both in peripheral and central retinal areas (not shown). Finally, Visomitin was shown to cause 3-fold reduction in the number of atrophic and dystrophic RPE loci adjacent to the areas of ONL damage and photoreceptor cell death (when compared to the control, Table 1). In general, according to quantitative assessment of the state of retina in the control and experimental samples of the posterior eye segment, the results of which are presented in Table 1, 3 h after illumination the state of retina remained close to intact in the vast majority of animals of the experimental group.
Table 1. Preventive effect of Visomitin
SkQ1-containing eye drops on photoinduced retinal changes in albino
rats
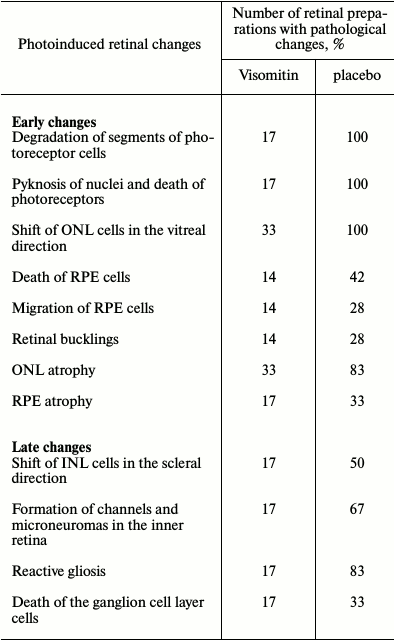
Note: Rats received Visomitin eye drops or similar drops without SkQ1
(placebo) for 15 days, and then were illuminated with bright light.
Photoinduced retinal changes were studied after 3 h (early changes) or
after 15 days (late changes), taking seven preparations from each group
for early changes and six preparations from each group for late
changes.
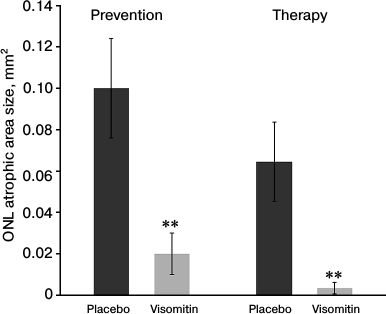
Fifteen days after illumination, ONL multifocal atrophy was quite pronounced, and so we compared atrophy in the areas of this layer in experimental and control retinal samples using two samples from each group of rats. The average area size of ONL atrophy in the experimental group of animals was found to be significantly smaller than in the control (Fig. 4), i.e. late changes in the outer neural retina were also sensitive to the protective effect of Visomitin. The data presented in Table 1 indicate that the same holds true for the late changes in RPE and inner retina. Areas of hypertrophy of the processes of Müller glial cells associated with the areas of ONL damage were present in the INL of the control samples (Fig. 3), whereas in the experimental samples reactive Müller gliosis was far less pronounced. This is also true for the cases of such severe changes as the development of channels and microneuromas observed in the majority of control, but not in experimental retinal samples (Table 1).
Fig. 4. Protective effect of Visomitin SkQ1-containing eye drops on photoinduced retinal changes in albino rats. When studying the preventive (left) and therapeutic (right) effects of Visomitin, we compared ONL atrophic areas 15 days after illumination of the rats with bright light.
Therapeutic effect of Visomitin SkQ1-containing eye drops on retina of albino rats exposed to illumination with bright light. In this series of the experiments, the rats were first illuminated with bright light, and then SkQ1-containing eye drops Visomitin (experimental group) or similar drops but without SkQ1 (control group) were administered for two weeks. After 15 days, we compared the state of the retina in these groups to determine whether Visomitin has a therapeutic effect on the retina after the illumination of animals. Table 2 summarizes the results of quantitative evaluation of the late pathological changes in retinal samples from the control and experimental animals demonstrating the pronounced protective effect of Visomitin on the state of both outer and inner retina after light-induced damage.
Table 2. Therapeutic effect of Visomitin
SkQ1-containing eye drops on photoinduced retinal changes in albino
rats
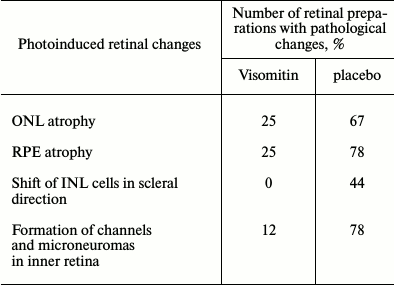
Note: Rats were first illuminated with bright light, and then received
Visomitin eye drops or similar drops without SkQ1 (placebo) for 15
days. Photoinduced retinal changes were studied immediately after the
course of Visomitin instillation 15 days after light-induced damage
(late changes) taking seven preparations from each group.
Measuring the areas of ONL atrophy in the experimental and control samples with the most pronounced changes (two samples from each group) showed that administration of Visomitin results in the significant reduction in the areas of ONL atrophy. Evaluation of the atrophic area size in this layer in experimental samples 15 days after illumination shows virtually complete absence of atrophic phenomena (Fig. 4). Visomitin also protects the inner segments of photoreceptor cells of the dorso-central retinal area, which serve as targets for light-induced damage from photoinduced shortening: the length of these segments in the experimental samples was 10 ± 1 µm, and in the control samples – 7 ± 1 µm. Visomitin caused three-fold greater preservation of RPE compared to the control samples (Table 2). According to these criteria, Visomitin has a pronounced therapeutic effect on the outer retina of rats, and its administration largely prevents cell death and local RPE destruction.
The inner retina of placebo-receiving rats demonstrated pronounced changes in cytoarchitectonics, including the formation of channels to drain the edema fluid and microneuromas, six times more often than the retina of experimental samples (Table 2). Thus, Visomitin-induced improvement of the state of the outer retina led to the reduction of pathological processes in the areas of the inner retina.
It should be added that solitary lymphocytic inflammatory infiltrates were found in the choroid of control, but not in the experimental samples of the eye posterior segment (not shown).
DISCUSSION
The LIRD (light-induced retinal degeneration) model is often used when studying light-dependent damages of the retinas of animals [17, 18]. According to this model, albino rat eyes are exposed to short-term illumination with light of high intensity (2000-3000 lx) or long-term illumination with light of low or medium intensity (200-500 lx) [20, 21]. Albino rats are used for the experiments because the light sensitivity of their retina is greater than that of pigmented animals due to the absence of melanin pigment in the albino eyes (melanin protects retinal structures against excessive light) [20, 22]. The uniqueness of the LIRD model is that in addition to the early damages in photoreceptor cells [24], it presents delayed damages in the RPE–Bruch’s membrane–choroid complex and even later compensatory-adaptive processes in the inner retinal layers [23]. Since intense illumination causes rather fast changes in retinal morphology, histological study of eye sections is commonly performed when using the LIRD model [17, 21, 23]. The degree of retinal damage in the different versions of the LIRD model varies and depends on a number of factors such as light intensity, exposure time, light source, as well as line, gender, age, and body temperature of the animals [17, 29]. For example, it is better to use female rats as their retinas are more sensitive to light-induced damage [29]. The dynamics of the destructive changes in the two major retinal cell populations serving as targets for light effects (photoreceptors and RPE cells) can also vary [30]. Numerous studies have been dedicated to research on molecular and cellular mechanisms that underlie the photoinduced damage of retinal cells [31] and the search for compounds that could prevent or at least reduce such damages. Since oxidative stress is an important component of the pathogenesis of photoinduced retinal damage, this search is mainly focused on compounds with antioxidant properties. For example, it was shown that curcumin and the green tea component epigallocatechin gallate, when used perorally, could reduce the risk of rat retina damage caused by bright light [32, 33]. Synthetic antioxidants such as TEMPOL-H can also protect photoreceptor cells, reducing the level of lipid peroxidation in them [34].
It has been demonstrated on a large number of in vitro and in vivo models that penetrating cation SkQ1 has high antioxidant activity [12-14]. However, the mechanism of SkQ1 effect on eye retina damaged by bright light remained unknown. We investigated this question in two series of experiments with the LIRD model [17, 18] using Visomitin SkQ1-containing eye drops. In the first series, when we studied the preventive effect of Visomitin on the retina, albino rats received Visomitin for two weeks, then they were illuminated with bright light and the morphology of their retina was analyzed 3 h and 15 days after the illumination. In the second series, when we studied the therapeutic effect of Visomitin on the retina, rats were first illuminated with bright light, then they received Visomitin for two weeks, and then the morphology of their retinas was analyzed. In both cases, the control animals received eye drops of the same composition but without SkQ1 following the same protocols.
Experiments on the preventive effect of Visomitin have shown (Table 1 and Figs. 2 and 4) that these eye drops have a pronounced protective effect on the albino rat retina, reducing the risk of the formation of early light-induced pathological changes and preventing their further development. Visomitin also inhibits the expression of late light-induced retinal changes, in particular, reactive gliosis, which increases the number and thickness of the processes of Müller glia cells. These results are consistent with data found in literature according to which proliferative activity of Müller glia cells and their specific density is increased in INL areas adjacent to outer retinal damages [35, 36]. Glia is known to be involved in the production and transport of a number of metabolites and neurotrophic factors in the retina [37]. It is also possible that retinal glial cells can be transformed into poorly differentiated precursors capable of differentiating into retinal neurons under certain conditions [38]. It can be assumed that retinal gliosis described in our experiments can serve as a compensatory event, and Visomitin-caused reduction in gliosis level reflects the lower level of photodamage.
According to the literature, light-induced damage of the retina (which is normally an immune-privileged zone) disturbs the blood–retinal barrier [39], resulting in the possibility of inflammatory infiltration of the retina [40]. In our experiments, illumination of the control rats led to the development of solitary lymphocytic infiltrates in the choroid of their retinas, which indicated damage to the blood–retinal barrier. Visomitin administration prevented the formation of such infiltrates, indicating the protective effect of the eye drops on this barrier.
Experiments studying the therapeutic effect of Visomitin have shown (Table 2 and Fig. 4) that these eye drops reduce the risk of the development of late light-induced pathological changes in the retina and lead to the preservation of the population of photoreceptor cells. Photoinduced death of these cells is known to occur not at the moment of illumination, but gradually over some time. During subsequent dark adaptation, photoreceptor cells start entering caspase-dependent apoptosis [41], leading to compensatory pathological changes in RPE [18]. The described therapeutic effect of Visomitin on the light-damaged retina can be explained by the protection of photoreceptor cells against apoptosis. In particular, the data obtained on cell cultures support this hypothesis. According to these data, SkQ and other mitochondria-addressed antioxidants prevent photoinduced apoptosis of HeLa cells by reducing the level of oxidative stress in them [42, 43]. It is also possible that the therapeutic effect of SkQ1 is connected with the restoration of the outer segments of some photoreceptor cells after their damage. For example, there is some evidence of the possibility of DNA repair in photoreceptor cells and reversibility of light-induced changes [44]. Moreover, restoration of outer photoreceptor segments in rats [45-47] and humans [48] is possible even in case of complete destruction of these photoreceptor segments as long as their perikaryon and cell nuclei remain intact. These observations suggest that Visomitin SkQ1-containing eye drops will be an effective medication to stimulate restoration mechanisms of retinal cells.
The described degenerative and atrophic light-induced retinal damages are typical for LIRD models in their nature and rate of development [17, 18, 20-24]. These damages were of multifocal character, being present in both peripheral and central retinal areas. The dorso-central retinal area was shown to be particularly prone to light-induced damage, which is consistent with its maximal sensitivity to light in rats and with data on topographic studies on the LIRD model [29, 49]. The dorso-central area of the rat retina can be considered as a functional analog of the human retinal macula, the area of sharpest vision. In this context, it is particularly important to note that etiology and mechanisms of animal retina damage under the conditions of the LIRD model have much in common with the mechanisms of the development of age-related retinal diseases. For this reason, the LIRD model is widely used in research on new approaches to prevention and treatment of age-related eye diseases [50], in particular, age-related macular degeneration, AMD [24, 33, 51], since constant exposure of the human retina to sunlight is considered to be one of the factors contributing to the development of AMD [4, 52, 53].
The data of the presented study indicate the ability of Visomitin SkQ1-containing eye drops to have pronounced preventive and therapeutic effects on the retina of rats exposed to bright light. Moreover, taking into account the previously demonstrated therapeutic effect of SkQ1 on AMD development in rapidly aging OXYS rats [54, 55], Visomitin can be considered as a potential medical preparation for the prevention and treatment of human retina damages, in particular, those accompanying age-related retinal macular degeneration.
The authors express their gratitude to Academician V. P. Skulachev for fruitful discussion of the results of this study.
This work was financially supported by the Research Institute of Mitoengineering, Lomonosov Moscow State University.
REFERENCES
1.Beckman, K. B., and Ames, B. N. (1998) The free
radical theory of aging matures, Physiol. Rev., 78,
547-581.
2.Skulachev, V. P. (2005) How to clean the dirtiest
place in the cell: cationic antioxidants as intramitochondrial ROS
scavengers, IUBMB Life, 57, 305-310.
3.Skulachev, V. P. (2006) Bioenergetic aspects of
apoptosis, necrosis and mitoptosis, Apoptosis, 11,
473-485.
4.Jarrett, S. G., and Boulton, M. E. (2012)
Consequences of oxidative stress in age-related macular degeneration,
Mol. Aspects Med., 33, 399-417.
5.Panfoli, I. (2012) Beneficial effect of
antioxidants in retinopathies: a new hypothesis, Med. Hypothesis
Discov. Innov. Ophthalmol., 1, 76-79.
6.Newell, F. W. (1992) Ophthalmology Principles
and Concepts (Mosby, C. V., ed.) 7th Edn., St. Louis.
7.Winkler, B. S., Boulton, M. E., Gottsch, J. D., and
Sternberg, P. (1999) Oxidative damage and age-related macular
degeneration, Mol. Vis., 5, 32.
8.Emerit, J., Edeas, M., and Bricaire, F. (2004)
Neurodegenerative diseases and oxidative stress, Biomed.
Pharmacother., 58, 39-46.
9.Roth, F., Bindewald, A., and Holz, F. G. (2004) Key
pathophysiological pathways in age-related macular disease, Graefes
Arch. Clin. Exp. Ophthalmol., 242, 710-716.
10.Kopitz, J., Holz, F. G., Kaemmerer, E., and
Schutt, F. (2004) Lipids and lipid peroxidation products in the
pathogenesis of age-related macular degeneration, Biochimie,
86, 825-831.
11.Tanito, M., Nishiyama, A., Tanaka, T., Masutani,
H., Nakamura, H., Yodoi, J., and Ohira, A. (2002) Change of redox
status and modulation by thiol replenishment in retinal photooxidative
damage, Invest. Ophthalmol. Vis. Sci., 43, 2392-2400.
12.Neroev, V. V., Archipova, M. M., Bakeeva, L. E.,
Fursova, A., Grigorian, E. N., Grishanova, A. Y., Iomdina, E. N.,
Ivashchenko, Zh. N., Katargina, L. A., Khoroshilova-Maslova, I. P.,
Kilina, O. V., Kolosova, N. G., Kopenkin, E. P., Korshunov, S. S.,
Kovaleva, N. A., Novikova, Y. P., Philippov, P. P., Pilipenko, D. I.,
Robustova, O. V., Saprunova, V. B., Senin, I. I., Skulachev, M. V.,
Sotnikova, L. F., Stefanova, N. A., Tikhomirova, N. K., Tsapenko, I.
V., Shchipanova, A. I., Zinovkin, R. A., and Skulachev, V. P. (2008)
Mitochondria-targeted plastoquinone derivatives as tools to interrupt
execution of the aging program. 4. Age-related eye disease. SkQ1
returns vision to blind animals, Biochemistry (Moscow),
73, 1317-1328.
13.Antonenko, Y. N., Avetisyan, A. V., Bakeeva, L.
E., Chernyak, B. V., Chertkov, V. A., Domnina, L. V., Ivanova, O. Y.,
Izyumov, D. S., Khailova, L. S., Klishin, S. S., Korshunova, G. A.,
Lyamzaev, K. G., Muntyan, M. S., Nepryakhina, O. K., Pashkovskaya, A.
A., Pletjushkina, O. Y., Pustovidko, A. V., Roginsky, V. A.,
Rokitskaya, T. I., Ruuge, E. K., Saprunova, V. B., Severina, I. I.,
Simonyan, R. A., Skulachev, I. V., Skulachev, M. V., Sumbatyan, N. V.,
Sviryaeva, I. V., Tashlitsky, V. N., Vassiliev, J. M., Vyssokikh, M.
Y., Yaguzhinsky, L. S., Zamyatnin, A. A., and Skulachev, V. P. (2008)
Mitochondria-targeted plastoquinone derivatives as tools to interrupt
execution of the aging program. 1. Cationic plastoquinone derivatives:
synthesis and in vitro studies, Biochemistry
(Moscow), 73, 1273-1287.
14.Agapova, L. S., Chernyak, B. V., Domnina, L. V.,
Dugina, V. B., Efimenko, A. Y., Fetisova, E. K., Ivanova, O. Y.,
Kalinina, N. I., Khromova, N. V., Kopnin, B. P., Kopnin, P. B.,
Korotetskaya, M. V., Lichinitser, M. R., Lukashev, A. L., Pletjushkina,
O. Y., Popova, E. N., Skulachev, M. V., Shagieva, G. S., Stepanova, E.
V., Titova, E. V., Tkachuk, V. A., Vasiliev, J. M., and Skulachev, V.
P. (2008) Mitochondria-targeted plastoquinone derivatives as tools to
interrupt execution of the aging program. 3. Inhibitory effect of SkQ1
on tumor development from p53-deficient cells, Biochemistry
(Moscow), 73, 1300-1316.
15.Grigoryan, E. N., Novikova, Y. P., Gancharova, O.
S., Kilina, O. V., and Philippov, P. P. (2012) New antioxidant SkQ1 is
an effective protector of rat eye retinal pigment epithelium and
choroid under conditions of long-term organotypic cultivation, Adv.
Aging Res., 1, 31-37.
16.Grigoryan, E., Novikova, Y., Kilina, O., and
Philippov, P. (2013) New antioxidant SkQ1 is an effective protector of
rat neural retina under conditions of long-term organotypic
cultivation, Adv. Aging Res., 2, 65-71.
17.Noell, W. K., Walker, V. S., Kang, B. S., and
Berman, S. (1966) Retinal damage by light in rats, Invest.
Ophthalmol., 5, 450-473.
18.Stone, J., Maslim, J., Valter-Kocsi, K., Mervin,
K., Bowers, F., Chu, Y., Barnett, N., Provis, J., Lewis, G., Fisher, S.
K., Bisti, S., Gargini, C., Cervetto, L., Merin, S., and Peer, J.
(1999) Mechanisms of photoreceptor death and survival in mammalian
retina, Prog. Retin. Eye Res., 18, 689-735.
19.Komarek, V., Gembardt, C., Krinke, A.-L.,
Mahrous, T., and Schaetti, P. (2000) Synopsis of the organ anatomy, in
The Laboratory Rat (Krinke, G., ed.) Academic Press, San Diego,
pp. 283-319.
20.Wasowicz, M., Morice, C., Ferrari, P., Callebert,
J., and Versaux-Botteri, C. (2002) Long-term effects of light damage on
the retina of albino and pigmented rats, Invest. Ophthalmol. Vis.
Sci., 43, 813-820.
21.Bennett, M. H., Dyer, R. F., and Dunn, J. D.
(1972) Light induced retinal degeneration: effect upon light-dark
discrimination, Exp. Neurol., 34, 434-445.
22.Williams, R. A., Howard, A. G., and Williams, T.
P. (1985) Retinal injury on pigmented and albino rats exposed to low
intensive cyclic light after unique mydriatic influence, Curr. Eye
Res., 4, 97-102.
23.Williams, T. P., and Howell, W. L. (1983) Action
spectrum of retinal light-damage in albino rats, Invest. Ophthalmol.
Vis. Sci., 24, 285-287.
24.Marc, R. E., Jones, B. W., Watt, C. B.,
Vazquez-Chona, F., Vaughan, D. K., and Organisciak, D. T. (2008)
Extreme retinal remodeling triggered by light damage: implications for
age related macular degeneration, Mol. Vis., 14,
782-806.
25.Abler, A. S., Chang, C. J., Ful, J., Tso, M. O.,
and Lam, T. T. (1996) Photic injury triggers apoptosis of photoreceptor
cells, Res. Commun. Mol. Pathol. Pharmacol., 92,
177-189.
26.Lin, Y., Jones, B. W., Liu, A., Vazquez-Chona, F.
R., Lauritzen, J. S., Ferrell, W. D., and Marc, R. E. (2012) Rapid
glutamate receptor 2 trafficking during retinal degeneration, Mol.
Neurodegener., 7, 7.
27.Kuwabara, T., and Gorn, R. A. (1968) Retinal
damage by visible light. An electron microscopic study, Arch.
Ophthalmol., 79, 69-78.
28.Schmidt, R. E., and Zuclich, J. A. (1980) Retinal
lesions due to ultraviolet laser exposure, Invest. Ophthalmol. Vis.
Sci., 19, 1166-1175.
29.De Vera Mudry, M. C., Kronenberg, S., Komatsu,
S., and Aguirre, G. D. (2013) Blinded by the light: retinal
phototoxicity in the context of safety studies, Toxicol.
Pathol., 41, 813-825.
30.Hafezi, F., Marti, A., Munz, K., and Reme, C. E.
(1997) Light-induced apoptosis: differential timing in the retina and
pigment epithelium, Exp. Eye Res., 64, 963-970.
31.Wenzel, A., Grimm, C., Samardzija, M., and Reme,
C. E. (2005) Molecular mechanisms of light-induced photoreceptor
apoptosis and neuroprotection for retinal degeneration, Prog. Retin.
Eye Res., 24, 275-306.
32.Costa, B. L., Fawcett, R., Li, G.-Y., Safa, R.,
and Osborne, N. N. (2008) Orally administered epigallocatechin gallate
attenuates light-induced photoreceptor damage, Brain Res. Bull.,
76, 412-423.
33.Mandal, M. N. A., Patlolla, J. M. R., Zheng, L.,
Agbaga, M.-P., Tran, J.-T. A., Wicker, L., Kasus-Jacobi, A., Elliott,
M. H., Rao, C. V., and Anderson, R. E. (2009) Curcumin protects retinal
cells from light- and oxidant stress-induced cell death, Free Radic.
Biol. Med., 46, 672-679.
34.Tanito, M., Li, F., Elliott, M. H., Dittmar, M.,
and Anderson, R. E. (2007) Protective effect of TEMPOL derivatives
against light-induced retinal damage in rats, Invest. Ophthalmol.
Vis. Sci., 48, 1900-1905.
35.Lewis, G. P., Guerin, C. J., Anderson, D. H.,
Matsumoto, B., and Fisher, S. K. (1994) Rapid changes in the expression
of glial cell proteins caused by experimental retinal detachment,
Am. J. Ophthalmol., 118, 368-376.
36.Fisher, S. K., Erickson, P. A., Lewis, G. P., and
Anderson, D. H. (1991) Intraretinal proliferation induced by retinal
detachment, Invest. Ophthalmol. Vis. Sci., 32,
1739-1748.
37.Bringmann, A., Pannicke, T., Grosche, J.,
Francke, M., Wiedemann, P., Skatchkov, S. N., Osborne, N. N., and
Reichenbach, A. (2006) Muller cells in the healthy and diseased retina,
Prog. Retin. Eye Res., 25, 397-424.
38.Jadhav, A. P., Roesch, K., and Cepko, C. L.
(2009) Development and neurogenic potential of Muller glial cells in
the vertebrate retina, Prog. Retin. Eye Res., 28,
249-262.
39.Cachafeiro, M., Bemelmans, A. P., Samardzija, M.,
Afanasieva, T., Pournaras, J. A., Grimm, C., Kostic, C., Philippe, S.,
Wenzel, A., and Arsenijevic, Y. (2013) Hyperactivation of retina by
light in mice leads to photoreceptor cell death mediated by VEGF and
retinal pigment epithelium permeability, Cell Death Dis.,
4, e781.
40.Greenwood, J. (1992) The blood-retinal barrier in
experimental autoimmune uveoretinitis (EAU): a review, Curr. Eye
Res, 11 (Suppl.), 25-32.
41.Perche, O., Doly, M., and Ranchon-Cole, I. (2007)
Caspase-dependent apoptosis in light-induced retinal degeneration,
Invest. Ophthalmol. Vis. Sci., 48, 2753-2759.
42.Antonenko, Y. N., Roginsky, V. A., Pashkovskaya,
A. A., Rokitskaya, T. I., Kotova, E. A., Zaspa, A. A., Chernyak, B. V.,
and Skulachev, V. P. (2008) Protective effects of mitochondria-targeted
antioxidant SkQ in aqueous and lipid membrane environments, J.
Membr. Biol., 222, 141-149.
43.Chernyak, B. V., Izyumov, D. S., Lyamzaev, K. G.,
Pashkovskaya, A. A., Pletjushkina, O. Y., Antonenko, Y. N., Sakharov,
D. V., Wirtz, K. W. A., and Skulachev, V. P. (2006) Production of
reactive oxygen species in mitochondria of HeLa cells under oxidative
stress, Biochim. Biophys. Acta, 1757, 525-534.
44.Gordon, W. C., Casey, D. M., Lukiw, W. J., and
Bazan, N. G. (2002) DNA damage and repair in light-induced
photoreceptor degeneration, Invest. Ophthalmol. Vis. Sci.,
43, 3511-3521.
45.Xia, X., Li, Y., Huang, D., Wang, Z., Luo, L.,
Song, Y., Zhao, L., and Wen, R. (2011) Oncostatin M protects rod
and cone photoreceptors and promotes regeneration of cone outer segment
in a rat model of retinal degeneration, PLoS One, 6,
e18282.
46.Wen, R., Tao, W., Luo, L., Huang, D., Kauper, K.,
Stabila, P., LaVail, M. M., Laties, A. M., and Li, Y. (2012)
Regeneration of cone outer segments induced by CNTF, Adv. Exp. Med.
Biol., 723, 93-99.
47.Youssef, P. N., Sheibani, N., and Albert, D. M.
(2011) Retinal light toxicity, Eye, 25, 1-14.
48.Forooghian, F., Stetson, P. F., Gross, N. E., and
Meyerle, C. B. (2010) Quantitative assessment of photoreceptor recovery
in atypical multiple evanescent white dot syndrome, Ophthalm. Surg.
Lasers Imag., 41 (Suppl.), 77-80.
49.Tanito, M., Kaidzu, S., Ohira, A., and Anderson,
R. E. (2008) Topography of retinal damage in light-exposed albino rats,
Exp. Eye Res., 87, 292-295.
50.Organisciak, D. T., Darrow, R. M., Rapp, C. M.,
Smuts, J. P., Armstrong, D. W., and Lang, J. C. (2013) Prevention of
retinal light damage by zinc oxide combined with rosemary extract,
Mol. Vis., 19, 1433-1445.
51.Ojino, K., Shimazawa, M., Ohno, Y., Otsuka, T.,
Tsuruma, K., and Hara, H. (2014) Protective effect of SUN N8075, a free
radical scavenger, against excessive light-induced retinal damage in
mice, Biol. Pharm. Bull., 37, 424-430.
52.Arnault, E., Barrau, C., Nanteau, C., Gondouin,
P., Bigot, K., Vienot, F., Gutman, E., Fontaine, V., Villette, T.,
Cohen-Tannoudji, D., Sahel, J.-A., and Picaud, S. (2013) Phototoxic
action spectrum on a retinal pigment epithelium model of age-related
macular degeneration exposed to sunlight normalized conditions, PLoS
One, 8, e71398.
53.Kernt, M., Thiele, S., Neubauer, A. S., Koenig,
S., Hirneiss, C., Haritoglou, C., Ulbig, M. W., and Kampik, A. (2012)
Inhibitory activity of ranibizumab, sorafenib, and pazopanib on
light-induced overexpression of platelet-derived growth factor and
vascular endothelial growth factor A and the vascular endothelial
growth factor A receptors 1 and 2 and neuropilin 1 and 2,
Retina, 32, 1652-1663.
54.Saprunova, V. B., Pilipenko, D. I., Alexeevsky,
A. V., Fursova, A. Z., Kolosova, N. G., and Bakeeva, L. E. (2010)
Lipofuscin granule dynamics during development of age-related macular
degeneration, Biochemistry (Moscow), 75, 130-138.
55.Markovets, A. M., Fursova, A. Z., and Kolosova,
N. G. (2011) Therapeutic action of the mitochondria-targeted
antioxidant SkQ1 on retinopathy in OXYS rats linked with improvement of
VEGF and PEDF gene expression, PLoS One, 6, e21682.